How to cite: Löser, 2022, A new coral family and three new genera (Scleractinia) from the Lower Cretaceous of Puebla and Sonora Mexico: Revista Mexicana de Ciencias Geológicas, v. 39, núm. 3, p. 220-229.
REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS
v. 39, núm. 3, 2022, p. 220-229
DOI: http://dx.doi.org/10.22201/cgeo.20072902e.2022.3.1698
A new coral family and three new genera (Scleractinia) from the Lower Cretaceous of Puebla and Sonora, Mexico
Hannes Löser
Universidad Nacional Autónoma de México, Instituto de Geología, Estación Regional del Noroeste, L.D.Colosio y Madrid s/n, Col. Los Arcos, 83110 Hermosillo, Sonora, Mexico.
loeser@paleotax.de
ABSTRACT
Solitary corals currently assigned to the genus Plesiosmilia are common elements in Late Jurassic to Early Cretaceous coral faunas. The genus itself is poorly defined and its systematic position is questionable. Moreover, examination of type material has shown that the genus lumps corals of different morphologies. Here, one common coral type is separated in a new solitary coral genus as well as two new phaceloid coral genera with a comparable morphology. The three new and another three existing genera are collected in a new family. The newly described material comes from the Upper Valanginian-Lower Hauterivian of San Juan Raya (Puebla) and the Lower Albian of Rayón (Sonora, both Mexico).
Key words: taxonomy; Hauterivian-Valanginian; systematics; fossil; Mexico; Puebla; Sonora.
RESUMEN
Corales solitarios actualmente asignados al género Plesiosmilia son comunes en faunas de corales del Jurásico Tardío y Cretácico Temprano. El género está mal definido y su posición sistemática es dudosa. Además, el examen de material tipo ha mostrado que el género incluye corales de diferentes morfologías. Aquí, un grupo común de corales es separado en un género nuevo de coral solitario y dos géneros nuevos de corales faceloides. Los tres géneros nuevos pertenecen a una familia nueva. El material descrito viene del Valangiano Superior-Hauteriviano Inferior del área de San Juan Raya (Puebla) y del Albiano Inferior de Rayón (Sonora, ambos México).
Palabras clave: corales; Cretácico; Scleractinia; México; Puebla; Sonora.
Manuscript received: june 26, 2022
Corrected manuscript received: october 4, 2022
Manuscript accepted: october 13, 2022
INTRODUCTION
Since Alloiteau (1952, 1957), the classification of the Mesozoic corals is mainly based on the microstructure of the septal blade, a principal morphologic part of the skeleton of Scleractinian corals. Alloiteau was followed by later authors (Morycowa, 1964, 1971; Roniewicz and Morycowa, 1993; Roniewicz, 1996, among many others). Septal microstructure refers principally to the size of the trabeculae, calcification centres that make up the septa. Whereas in earlier studies the absolute size of trabeculae was taken into account (Roniewicz, 1996), Löser (2016) proposed to relate the size of the trabeculae to the thickness of septa. Moreover, it was shown that the microstructure may vary within the same coral (Löser, 2016: fig. 3.3.1.4) depending on its position in the septal blade. In addition to the septal microstructure, the presence of perforations in the septa, and the presence or absence of synapticulae, and other peculiar characteristics such as pennulae or auriculae, are considered crucial for the definition of the families and higher taxonomic ranks such as superfamilies or suborders. The way in which to classify Mesozoic corals – based on the fine structure of the skeleton, as proposed by Alloiteau and refined by later authors – works properly when these skeletal structures are preserved. The skeleton of Scleractinian corals is made of aragonite, a mineralogical variation of calcite that is usually post-mortem diagenetically transformed into calcite. During this process, skeletal microstructures can become altered or completely lost. A second diagenesis may replace the calcite by silica, resulting in even more alteration. Hence, the preservation of microstructures is an exception, not the rule. Of course, it is possible to compare material that has no microstructures at all to a genus that is based on a proper study of the septal microstructure. But it is not possible to form a diagnosis for a new genus on material without showing septal microstructures, because the systematic position cannot be proven without its knowledge.
In the 19th century, post-palaeozoic corals were principally classified into families. This concept was improved by Americans Vaughan and Wells (1943) who introduced suborders. Their French counterpart, James Alloiteau and later authors followed them and applied suborders within the order Scleractinia for the distinction of larger groups. Nevertheless, only some suborders are well limited or clearly based on type material (Löser, 2016), whereas others are poorly defined and/or used in a very broad sense. Because the order group is not covered by the code of the International Commission of Zoological Nomenclature (ICZN) it makes this taxonomic level highly conceptual. In order to avoid conceptual and non-type-based classification on the one hand, and to present a more consistent concept for the taxonomic level above the family, on the other, Löser (2016) introduced the superfamily as a principal taxonomic level to group Mesozoic Scleractinian corals, distinguishing 27 superfamilies and 56 families.
Some superfamilies and families had an informal character because no formally established names were available. One of these groups is that of the genus Plesiosmilia Milaschewitsch, 1876, and related or synonymous genera first introduced in Löser (2013a). Plesiosmilia itself cannot serve as type for a new family because the type of the type species Plesiosmilia turbinata Milaschewitsch, 1876 is silicified. The types of the type species of possible junior synonyms (Elasmosmilia Beauvais, 1961, Lophosmilia Milne Edwards and Haime, 1848a, Peplosmilia Milne Edwards and Haime, 1851, among others, see below) are equally poorly preserved. There exists the family Axosmiliidae Geyer, 1955 based on Axosmilia Milne Edwards and Haime, 1848a, but the type specimen of Caryophyllia extinctorium Michelin, 1841 (type species of Axosmilia) is equally extremely poorly preserved. The same applies to the family Ellipsosmiliidae Alloiteau, 1957 where the type specimen of Montlivaltia cornucopia Milne Edwards and Haime, 1848b, type species of Ellipsosmilia d’Orbigny, 1849, is not available and, moreover, from a locality where all corals are silicified.
Not only is there Plesiosmilia in this group, there are also more solitary (Actinosmilia d’Orbigny, 1849, Carantoseris Alloiteau 1952, Cenomanosmilia Alloiteau 1949, Ceratosmilia Alloiteau, 1957, Dautlusmilia Kuzmicheva, 2002, Plesiolites Löser, Steuber and Löser, 2018, Trochophyllia Alloiteau, 1952) and yet unnamed phaceloid corals. All these genera cannot be assigned with certainty to this informal group because it is poorly defined. The Plesiosmilia Group is widely distributed in space and time ranging from the Jurassic into the Late Cretaceous. The present contribution therefore introduces a new formal genus and family name in order to replace the Plesiosmilia Group by a more proper unit. Moreover, another two related genera are established.
Figure 1. Study area, general view.
Study area
The material described herein comes from two localities in Mexico (Figure 1). A part of the material comes from the famous outcrop area of San Ruan Raya in Puebla (Figure 2a, 2b). Coral occurrence, localities and stratigraphy were discussed by Löser et al. (2013). For a long period of time this area was considered to be Aptian in age, until the study of nano-fossils that revealed an Upper Valanginian to Lower Hauterivian age (González León et al., 2015), which is supported by the composition of the coral fauna (Löser, 2021). The studied material comes from the historic Aguilar collection, which is kept in the fossil collection of the Institute of Geology at the UNAM (National Autonomous University of Mexico) in Mexico City. The exact locality of the material is unknown. The other part of the material comes from central Sonora (Figure 2c, 2d). Around the village of Cerro de Oro, shallow marine strata of the Cerro de Oro Fm (Bisbee Group) yields fossils with an Upper Barremian to Lower Aptian age (González León and Lucas, 1995; Baron-Szabo and González León, 1999; Löser, 2011; Velázquez-Heras et al., 2021). The studied material is kept at the Northwest Regional Station (ERNO) in Hermosillo, Sonora. Because the studied material comes from collections, no exact sample positions can be given.
Figure 2. Study area. a, b, Puebla. c, d, Sonora.
MATERIAL AND METHODS
For the present study, approximately 300 coral specimens that are similar to Plesiosmilia ranging from the Upper Jurassic to the Upper Cretaceous were examined, and 75 of them are type specimens. Around 125 thin sections were available for the study. Thin sections were scanned by passing light through them using a flatbed scanner with an optical resolution of 6,400 dpi. Scanned images were then transferred to greyscale bit maps. Their quality was amended using histogram contrast manipulation (contrast stretching) where possible. Corallite dimensions of each specimen were systematically measured. In the phaceloid corals, the largest number of possible measurements was taken. This number was mainly determined by the size and quality of the thin section and the size of the single corallites, in relation to the size of the thin sections. Septa were counted for numerous corallites. For the corallite diameter and septal counts in one thin section of the colonial corals, the following values were obtained:
n, number of measurements or counts
min–max, lowest and highest measured or counted values (mm for measurements)
µ, arithmetic mean (average)
s, standard deviation
v, coefficient of variation according to K. Pearson
µ±s, first interval
c, corallite diameter
cmin, small corallite diameter
cmax, Large corallite diameter
s, septa.
Thin sections were measured and values were calculated using the Palaeontological Database System PaleoTax, module PaleoTax/Measure (https://www.paleotax.de/measure). Morphometric data of the corals were compared against those of specimens in worldwide fossil coral collections, and an associated image database located in the ERNO. Data storage and processing were carried out using the PaleoTax database programme (Löser, 2004). Collection acronyms as follows: ERNO, Northwest Regional Station at the UNAM (Hermosillo, Sonora, Mexico); IGM, Institute of Geology at the UNAM, Mexico City, Mexico; MGB, Geological Museum of the City of Barcelona (Catalonia, Spain).
SYSTEMATIC DESCRIPTION
Order Scleractinia Bourne, 1900
Superfamily Misistelloidea Eliášová, 1976
Diagnosis. Solitary and colonial (astreoid, phaceloid, plocoid) corals. Septa compact, in varying thickness, in a subregular radial symmetry. Septal outline enlarged rhombic. Septa not connected to each other (Rayasmiliidae) or only in the centre of the corallite (Misistellidae). Lateral faces with fine granulae or smooth, upper margin smooth. Lonsdaleoid and main septa absent. Microstructure of septa of small trabeculae. Synapticulae absent. Pali absent, columella present in most genera, lamellar or by septal fusion. Endotheca present. Marginarium absent. Wall absent or epithecal. No coenosteum. Budding intracalicinal.
Families
Misistellidae Eliášová, 1976 and Rayasmiliidae n. fam. (formerly Plesiosmilia group).
Family Rayasmiliidae new family
Type genus. Rayasmilia n. gen.
Zoobank ID. urn:lsid:zoobank.org:act:EB179F35-6BA4-4263-805E-9413199D27BE.
Diagnosis. Solitary and phaceloid corals. Septa compact, in varying thickness, in a regular or subregular radial symmetry. Septal outline enlarged rhombic (bicuneiforme in French). Septa not connected to each other. Lateral faces with fine granulae or smooth, upper margin smooth. Lonsdaleoid and main septa absent. Microstructure of septa of small trabeculae. Synapticulae absent. Pali absent, a lamellar columella is present in some genera. Endotheca present. Marginarium absent. Wall absent or epithecal. No coenosteum. Budding intracalicinal.
Comparison. The new family differs in two aspects from the Misistellidae. The septa in the Misistellidae shows a zigzag pattern towards the corallite centre. They are connected in the corallite centre to form a type of columella, and they are also connected to each other.
Discussion. In Löser (2013a) the family was introduced as the informal Plesiosmilia-group. The genus Plesiosmilia cannot serve as a type taxon, since its type and topotypical material is silicified and fine skeletal structures are, therefore, unknown. The family belongs to the superfamily Misistelloidea that is based on the family Misistellidae. The family was originally assigned to the suborder Astraeoida where it is misplaced. The suborder Astraeoida Alloiteau, 1952 was originally characterised as having compact septa with a strongly ornamented upper septal border. The latter may imply that the trabeculae are large. Later, the suborder was synonymised with the suborder Faviina Vaughan and Wells, 1943 that has clearly large trabeculae (Garberoglio et al., 2021). The type of the type species of the name-giving genus Astrea Lamarck, 1801 (type species Madrepora rotulosa Ellis and Solander, 1786) was never the object of a detailed examination (the type is figured in Löser 2016). Further, the new family cannot be assigned to the suborder Meandrinina Alloiteau, 1952 that has – in comparison to the Astraeoida and Faviina – smooth distal margins; however, the septal blades are made up of medium-sized trabeculae (Löser, 2016, fig. 3.3.1.3). For those coral workers who still apply suborders, the new family remains without suborder.
Genera
For the moment, we include here six genera (Table 1). The number of genera is surely higher but since many genera lack information about their fine skeletal structure, mainly the septal microstructure, they probably cannot be assigned to any family. Principally for the genus Rayasmilia, many similar genera exist (see below for more details). Morycowa and Marcopoulou-Diacantoni (2002) illustrate under the name Plesiofavia dubia (de Fromentel, 1857) astreoid colonies that have the microstructures as stated for the family. This material cannot be assigned to Plesiofavia because this genus belongs as a junior synonym of Ovalastrea d’Orbigny, 1849 to the family Latomeandridae de Fromentel, 1861 (see Löser, 2016).
Table 1. Genera of the Rayasmiliidae n. fam.
|
Phaceloid |
With columella |
Sonoraphyllia |
|
|
Without columella |
Rayaphyllia |
||
|
Solitary |
Cyclotitid |
Plesiolites |
|
|
Turbinate |
Columella lamellar |
Rayasmilia |
|
|
Columella spongiosa |
Dautlusmilia |
||
|
Without columella |
Trochophylia |
||
Distribution
Upper Jurassic to Cretaceous, worldwide. It is possible that the family already occurs in the Middle Jurassic, but no sectioned material was to hand to confirm this. Further, the family may continue into the Palaeogene, but Cenozoic corals are usually not examined using thin sections and their microstructure is, therefore, unknown.
Genus Rayasmilia new genus
Type species. Rayasmilia salvata n. sp.
Zoobank ID. urn:lsid:zoobank.org:act:1A96D599-5E3E-42F7-813A-DBA8ADC6725C.
Diagnosis. Solitary turbinate coral of circular to elliptical outline. Septa compact, made of small trabeculae, not connected to each other, in a regular radial symmetry. Wall absent. Epitheca present. Endotheca well developed. Columella lamellar.
Description. Solitary turbinate coral. Corallite outline circular to elliptical, diameter up to 45 mm, but generally below 30 mm, pit depressed. Septa compact. Microstructure of small-sized trabeculae, septa with a median dark line. Septa in cross section bicuneiform thich close to the wall, thinner towards the centre. Septa strong with a thickness of one millimetre or more. Symmetry of septa radial and regularly in varying systems, but 12 being most common. Cycles of septa regular. Septal cycles differ in length and thickness. First septal cycle extends to the corallite centre, later cycles are subsequently shorter. Septa are not connected to each other. Septal upper margin smooth, lateral face smooth or finely granulated, inner margin smooth. Pali absent. Some septa of the first cycle may, rarely, be attached to the columella. Costae present but short, smooth on their surface. Synapticulae absent. Columella lamellar. Endotheca consists of numerous dissepiments. Wall absent. Epitheca present.
Etymology. After the type locality San Juan Raya. The Spanish word raya translates into line, corresponding well to the microstructural feature of the new genus. The suffix -smilia is commonly used for solitary corals.
Occurrence. Upper Jurassic to Cenomanian, worldwide.
Comparison. The new genus is difficult to compare to similar coral genera because most of the type material of these genera does not show microstructures. The following genera could be related to the new genus:
Axiphyllum Quenstedt, 1880. – The type material of the type species is silicified. The genus is considered to be a synonym of
Plesiosmilia.
Axosmilia. – The type of the type species is poorly preserved and there are no thin sections available.
Cenomanosmilia. – There are no thin sections available and topotypical material could not be obtained since the type locality is no longer accessible. The types seem to be strongly recrystallised.
Ceratosmilia. – The septal microstructure is unknown because the type of the type species is silicified.
Dautlusmilia. – The columella is stronger and seems to be spongiose.
Elasmosmilia. – The type of the type species is completely recrystallised.
Ellipsosmilia. – The type of the type species is silicified.
Lophosmilia. – The type material of the type species is lost. There exists only an unjustified neotype. There are no thin sections
available.
Peplosmilia. – The type of the type species is only fragmentarily preserved.
Plesiolites. – The genus is similar to Rayasmilia but has a cupolate form comparable to Cyclolites Lamarck, 1801.
Plesiosmilia. – The septal microstructure is unknown because the type of the type species is silicified.
Saltocyathus Morycowa and Masse, 1998. – The type of the type species is poorly preserved, the thin sections are fragmentary and show a strongly recrystallised coral.
Trochophyllia. – There are no thin sections available. The septal microstructure is unknown. The genus does not show a columella.
The genera Kobyphyllia Baron-Szabo and Fernández Mendiola, 1997 and Montlivaltia Lamouroux, 1821 are, in their outer appearance, very similar to the new genus, but they differ in larger trabeculae (easily visible at the upper and outer septal margin) and – in Montlivaltia – the absence of a columella. Material described as ?Placosmilia Milne Edwards and Haime, 1848a and ?Lasmogyra d’Orbigny, 1849 by Löser et al. (2019) is very similar but has larger trabeculae that are only visible in well preserved material. Caryophyllids have a very reduced endotheca and a septothecal wall. The genus Aulosmilia Alloiteau, 1952 is comparable in its general structure, but differs by way of its septal microstructure, septal outline, the T-shaped inner septal margins, a septothecal wall, and septa of younger cycles that are attached to septa of older cycles. Aulosmilia has its mayor distribution in the Upper Cretaceous and Palaeogene, but also rarely occurs in the Lower Cretaceous (for instance, Peplosmilia fromenteli Angelis d’Ossat, 1905 in Baron-Szabo and Steuber, 1996). Species with very thin septa (such as Axosmilia bella Reig Oriol, 1991, Axosmilia mexicana Wells, 1946, Pleurosmilia whitneyi Wells, 1933, or syntype MGB 20521 of Peplosmilia casanasi Angelis d'Ossat, 1905) are not included in Rayasmilia, at the present time; however, they may belong to Bathmosmilia Alloiteau, 1958. The same applies to similar solitary corals from the Upper Cretaceous (for instance, material assessed as Plesiosmilia in Löser, Steuber and Löser, 2018), whereby the microstructure consists of small trabeculae marked by a dark line that is traversed by perpendicular dark lines.
Species. It is only possible to confirm which species that were described under the above given long list of genera belonging to Rayasmilia, when the type material is available and a section is provided. To date, 85 species are assigned to the genus Plesiosmilia, 39 from the Jurassic, and 46 from the Cretaceous. The following species can be assigned with certainty to the genus Rayasmilia: Axosmilia almerai Angelis d’Ossat, 1905, Axosmilia angelisi Reig Oriol, 1991, Axosmilia bangoinensis Liao and Xia, 1985, Pleurosmilia barottei de Fromentel, 1867, Thecosmilia bassanii Prever, 1909a, Aulophyllia belbekensis Kuzmicheva, 2002, Axosmilia bofilli Angelis d’Ossat, 1905, Lophosmilia cenomana Alloiteau, 1949, Peplosmilia coquandi Angelis d’Ossat, 1905, Placosmilia crassisepta Prever, 1909a, Saltocyathus cumburindioensis Filkorn and Pantoja-Alor, 2015, Peplosmilia fromenteli Angelis d’Ossat, 1905, Coelosmilia gracilis Prever, 1909a, Pleurosmilia hennigi Dietrich, 1926, Pleurosmilia hidesimaensis Eguchi, 1951, Peplosmilia iberica Angelis d’Ossat, 1905, Peplosmilia infundibuliformis Alloiteau, 1936, Pleurosmilia neocomiensis de Fromentel, 1867, Pleurosmilia quaylei Wells, 1933, Pleurosmilia saxifisi Wells, 1933, Pleurosmilia schiosensis Boehm, 1897, Sphenotrochus straini Turnšek, LeMone and Scott, 2003, Axosmilia styphelos Liao and Xia, 1985, Rhipidosmilia tauridae Babaev and Krasnov, 1977, Peplosmilia thildae Angelis d'Ossat, 1905, Plesiosmilia tombecki Alloiteau, 1936, Saltocyathus urgonensis Morycowa and Masse, 1998, Pleurosmilia vaughani Angelis d’Ossat, 1905, Axosmilia viai Reig Oriol, 1991. The list encompasses only Cretaceous species.
Most common is a duodecameral symmetry, but there are also species with a decameral or hexameral septal symmetry. The number of septa does not increase with the growth of the coral. The septa grow in a very early ontogenetic stage of the coral. When this stage is reached, the coral mostly increase in height, but very little in diameter. Therefore the species can be distinguished by the septal symmetry, the number of septal cycles, and the diameter.
Rayasmilia salvata new species Figure 3
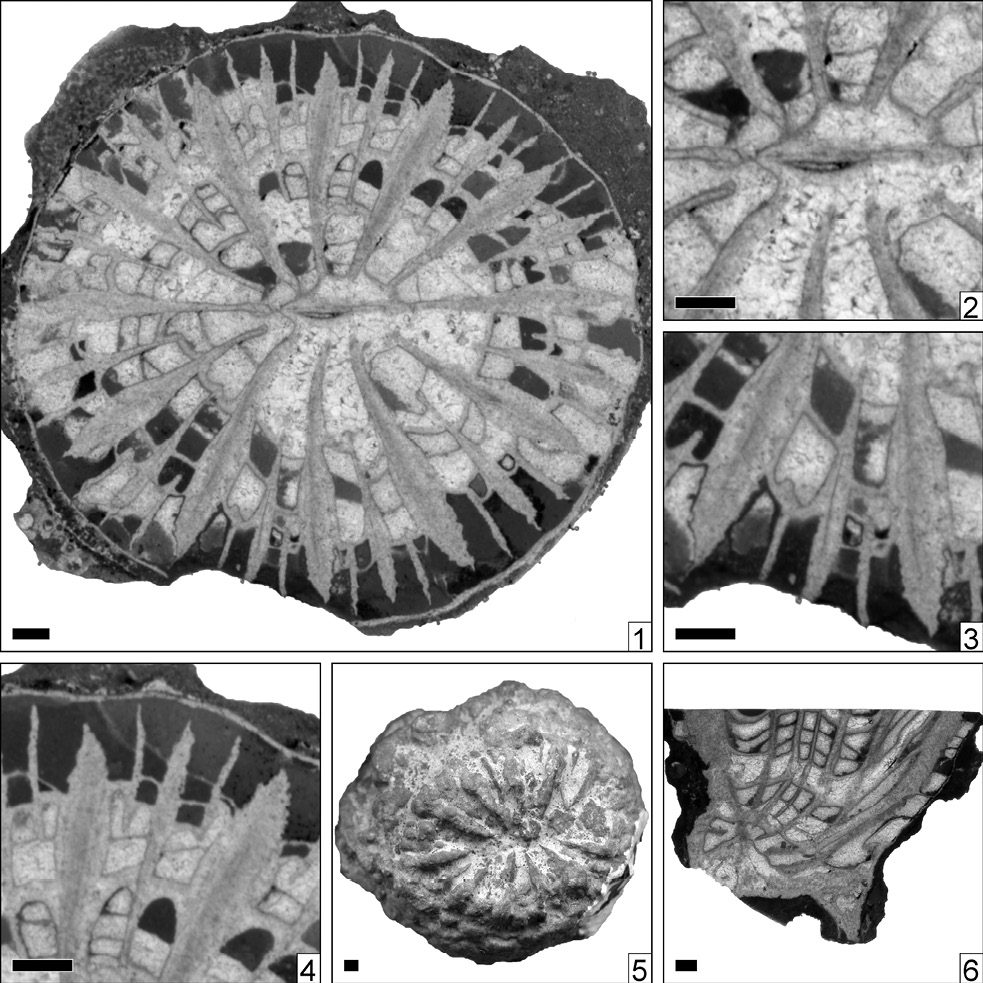
Figure 3. Rayasmilia salvata n. sp., n. gen. Holotype IGM 9193. 1, transversal thin section; 2, transversal thin section, detail of the corallite centre; 3, transversal thin section, detail of the microstructure; 4, transversal thin section, detail of the epitheca; 5, oral view; 6, longitudinal thin section. Scale bars = 1 mm.
v2013a Plesiosmilia hennigi (Dietrich, 1926) – Löser, fig. 5.10
v2013b Plesiosmilia hennigi (Dietrich, 1926) – Löser, fig. 3.1
Zoobank ID. urn:lsid:zoobank.org:act:CA986162-0B00-4F6C-8264-AABF29E961C4.
Diagnosis. Rayasmilia with a duodecameral septal symmetry with three septal cycles resulting in 48 septa. Corallite diameter about 16 mm.
Description. As for the genus.
Etymology. Feminine form of the Latin participle salvatus, to save. It refers to the intention to save or to assure the informal concept in the form of a better defined taxon.
Type. Holotype IGM 9193. Mexico, Puebla, San Juan Raya area. Cretaceous, San Juan Raya Formation, Upper Valanginian to Lower Hauterivian.
Measurements.
c 15.8 × 16.7 mm
s 12 s1 + 12 s2 + 24 s3
Occurrence. Based on available specimens, Berriasian to Aptian, worldwide.
Genus Rayaphyllia new genus
Type species. Rayaphyllia atheca n. sp.
Zoobank ID. urn:lsid:zoobank.org:act:4F03F810-D1FE-496F-8775-EA32A4330593.
Diagnosis. Phaceloid coral. Septa compact, made of small trabeculae, not connected to each other, in a subregular radial symmetry. Wall absent. Epitheca present. Endotheca well developed. Columella absent.
Description. Phaceloid colony. Corallite outline circular or elliptical, diameter up to 16 mm but generally below 10 mm. Septa compact. Microstructure of small-sized trabeculae, septa with a median dark line. Septa in cross-section, bicuneiform close to the wall, thinner towards the centre. Septal maximum thickness around 500 µm for the first cycle. Cycles of septa subregular. Septal cycles differ in length and thickness. First septal cycle extends to the calicular centre, later cycles are subsequently shorter. Septa occasionally connected, only by means of dissepiments. Septal distal margin unknown, lateral face and inner margin smooth. Pali absent. Costae hardly present, surface unknown. Synapticulae absent. Columella absent. Endotheca consists of dissepiments, in two regular rings in the type species. Wall absent. Epitheca present. Coenosteum absent. Budding intracalicinal.
Etymology. After the type locality San Juan Raya. The suffix -phyllia (Greek for leaf) is commonly used for phaceloid corals.
Occurrence. Aptian to Santonian, worldwide.
Discussion. The only comparable genus is Montlivaltoides He and Xiao, 1990, but the type material of the type species is poorly preserved and insufficiently described.
Species. The type species, Thecosmilia distefanoi Prever, 1909a, and with certain reservation Montlivaltoides ngariensis He and Xiao, 1990.
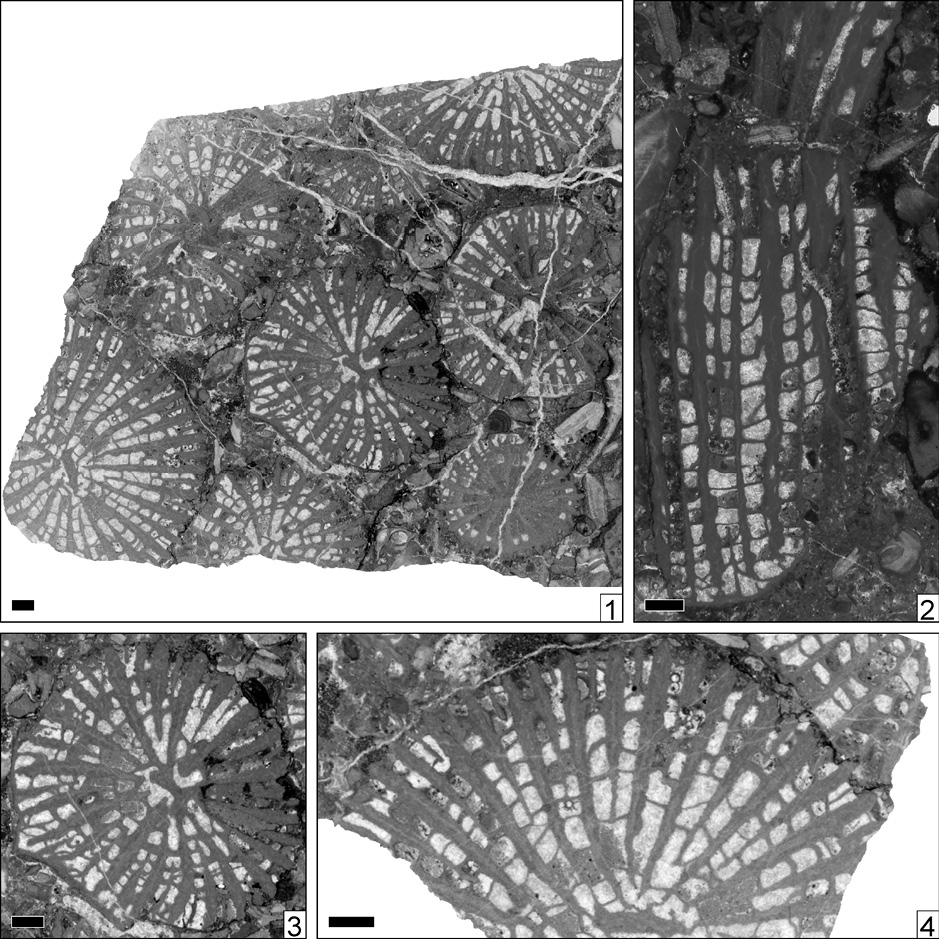
Rayaphyllia atheca new speciesFigure 4

Figure 4. Rayaphyllia atheca n. sp., n. gen. Holotype IGM 9204. 1, transversal thin section; 2-3, transversal thin section, detail; 4, longitudinal thin section. Scale bars = 1 mm.
v1889 Lophosmilia (?) spec. – Toula, p. 84, pl. 6, fig. 3
v2019 Montlivaltoides typicus He & Xiao, 1990 – Löser et al., p. 215, fig. 337
Zoobank ID. urn:lsid:zoobank.org:act:5FBE5E6F-F2E8-4F37-9395-02D9999A72ED.
Diagnosis. Rayaphyllia with a corallite diameter of six to eight millimetres, and between 34 and 48 septa.
Description. As for the genus.
Etymology. The Latin word theca (box, capsule, cover, container) refers in extant corals to the lateral boundary of the corallite. In Mesozoic corals, the term wall is more common. Atheca refers to the absence of a wall.
Type. Holotype IGM 9204. Mexico, Puebla, San Juan Raya area. Cretaceous, San Juan Raya Formation, Upper Valanginian to Lower Hauterivian.
Measurements.
|
n |
min-max |
µ |
s |
cv |
µ±s |
|
|
cmax |
7 |
6.92-8.66 |
7.94 |
0.67 |
8.5 |
7.26-8.61 |
|
cmin |
7 |
5.98-9.0 |
7.56 |
1.05 |
13.9 |
6.50-8.61 |
|
septa |
8 |
34-48 |
41.75 |
4.68 |
11.2 |
37-46 |
Occurrence. Valanginian to Santonian, worldwide.
Genus Sonoraphyllia new genus
Type species. Sonoraphyllia aurea n. sp.
Zoobank ID. urn:lsid:zoobank.org:act:F5137836-090A-4140-8EA1-EBDCAA3AAB43.
Diagnosis. Phaceloid coral. Septa compact, made of small trabeculae, not connected to each other, in a regular radial symmetry. Wall absent. Epitheca present. Endotheca well developed. Columella lamellar.
Description. Phaceloid colony. Calicular outline circular to elliptical, diameter 8-10 mm. Septa compact. Microstructure of small-sized trabeculae, septa with a median dark line. Septa in cross-section, bicuneiform close to the wall, thinner towards the centre. Septal maximum thickness 500 µm. Symmetry of septa irregularly radial with subregular cycles. Septal cycles differ in length and thickness. First septal cycle extends to the calicular centre, later cycles are subsequently shorter. Septa occasionally connected, only by means of dissepiments. Septal distal margin unknown, lateral face smooth, inner margin smooth. Pali absent. Some septa of the first cycle are attached to the columella. Costae hardly present, non-confluent, surface unknown. Synapticulae absent. Columella lamellar. Endotheca consists of dissepiments. Wall absent. Epitheca present. Coenosteum absent. Budding intracalicinal.
Etymology. Named after its origin, the Mexican state Sonora. The suffix -phyllia (Greek for leaf) is commonly used for phaceloid corals.
Occurrence. Tithonian to Middle Albian (?Cenomanian), worldwide.
Discussion. The only comparable genus is Montlivaltoides He and Xiao, 1990, but the type material of the type species is poorly preserved and insufficiently described.
Species. The type species, Aplophyllia marini Bataller, 1947, Aplosmilia tolmachoffana Wells, 1932, Placophyllia baingoingensis Wang, Sun, Wang, Zheng, Yue and Liao, 2020, Placophyllia bandeli Baron-Szabo, 1998, Placophyllia florosa Eliášová, 1976, and Thecosmilia hideshimaensis Eguchi, 1951. Material described by Prever (1909b) as Thecosmilia virgulina (Étallon, 1864) from the Tithonian of L’Aquila (Italy) may belong to this genus. There are more, but not yet formally established, species.
Sonoraphyllia aurea new speciesFigure 5
Figure 5. Sonoraphyllia aurea n. sp., n. gen. Holotype ERNO 2161. 1, transversal thin section; 2, longitudinal thin section; 3-4, transversal thin section, detail. Scale bars = 1 mm.
v1999 Placophyllia bandeli Baron-Szabo, 1998 – Baron-Szabo and González-León, p. 475, fig. 3e, f
Zoobank ID. urn:lsid:zoobank.org:act:FCBCD8F0-390A-45DF-A71D-3A3717E23BB0
Diagnosis. Sonoraphyllia with a corallite diameter of 8 to 10 mm, and 37 to 48 septa.
Description. As for the genus.
Etymology. Latin aurea for gold after the type locality, Cerro de Oro (Gold Hill).
Type. Holotype ERNO 2161 Mexico, Sonora, Cerro de Oro. Cretaceous, Cerro de Oro Formation, Upper Barremian to Lower Aptian.
Measurements.
cmin 7.5-8.2 mm
cmax 9.1-10.5 mm
septa 37-49
Occurrence. Upper Barremian to Middle Albian, worldwide.
DISCUSSION
Taxonomy
Before the fundamental works of Alloiteau became published, already more than 100 families within the order Scleractinia were established. Many of them are based on genera where the type material of the corresponding type species is now lost, poorly preserved or from localities where all corals are silicified (such as the Kimmeridgian Nattheim in southern Germany or the Cenomanian Isle de Aix area in western France). Numerous families are now considered to be synonymous. To date, more than 200 families (for less than 2,000 genera from the Triassic to recent) are formally established. This high number is due to a change in the classification concept that was first introduced by Alloiteau. The application of thin sections and the study of microscopic characteristics of the septal morphology created a more stable and clear concept. Many genera are still without a proper family and many families are still applied in a traditional concept based on macromorphological characteristics.
Palaeoecology
Whereas members of the phaceloid genera Sonoraphyllia and Rayaphyllia are rare, or not yet fully recognised in the fossil record, the genus Rayasmilia is rather common. Among all of the approximately 400 coral genera that occur in the Cretaceous, the genus Rayasmilia belongs to the occasional genera having nearly 60, out of 2,000 localities. Rayasmilia is more common during the Aptian and Albian. It occurs together with hermatypic colonial corals and, therefore, lived in shallow marine environments. Among the solitary corals that occur in the time period Hauterivian to Albian, Rayasmilia is the most common of all solitary corals. From the Cenomanian on, with increasing diversity of ahermatypic corals, the ahermatypic corals Micrabacia Milne Edwards and Haime, 1849 and Parasmilia Milne Edwards and Haime, 1848a are the most common solitary corals, that are not sharing the area with shallow marine colonial corals (Aulosmilia and Cyclolites being the most abundant solitary coral genera living together with hermatypic corals from the Cenomanian on). In Berriasian to Valanginian strata, the most common solitary coral is Montlivaltia. Rayasmilia has replaced Montlivaltia from the Hauterivian on, and was itself replaced by Aulosmilia and Cyclolites (for abundance calculation see Löser 2016: 113). This replacement process was gradual (Figure 6) with the exception of the Cenomanian/Turonian boundary where, during a short period, no coral data are available. Montlivaltia and Rayasmilia became extinct by the end of the Cenomanian, thus making Aulosmilia the dominating hermatypic turbinate coral.
Cyclolites cannot be compared to these three turbinate genera. It is a cupolate coral that laid unattached on the sea floor (currently, as with members of the genus Fungia Lamarck, 1801), whereas the turbinate corals were probably attached.
In shallow marine tropical environments, solitary corals are only presented by few genera (Fungia and allied genera, Scolymia Haime, 1852) where Fungia and allied genera practically do not live together with colonial corals (see, for instance, Hoeksema, 1989 ). The greater success of solitary corals in the Cretaceous can be explained by the lower degree of competition. Because the Cretaceous calcite sea was unfavourable to corals and calcarous algae, coral reef formation is almost unknown from the Cretaceous. The coral colonies remained small, rarely reaching more than 20 cm in diameter. In such an environment, the slower growing solitary corals could survive.

Figure 6. Proportion of the three genera Montlivaltia, Rayasmilia, and Aulosmilia through the Cretaceous, based on positively identified specimens. Specimens from localities with a range in time of more than 13.3 ma (duration of the longest stage, the Albian) are excluded. Number of included localities: Montlivaltia (42), Rayasmilia (45), and Aulosmilia (51). No data from the Early Turonian due to a global coral gap.
ACKNOWLEDGMENTS
This publication profited from funding by CONACyT project 52442-Q. Preparation of thin sections in the ERNO laboratory by Aimé Orcí (Hermosillo, Sonora, Mexico) as well in the thin section laboratory of the Geological Institute (UNAM, City of Mexico) is gratefully acknowledged. Reviews of Andrés López Pérez (Mexico City) and Ricardo Garberoglio (Buenos Aires) helped to improve the manuscript. A grammatical check was carried out by Matthew Copley (Barcelona).
REFERENCES
Alloiteau, J., 1936, Sur la présence, dans le calcaire a Spatangues de la Haute-Marne, de Plesiosmilia villersensis Koby: Bulletin de la Société géologique de France, (5)6, 507-510.
Alloiteau, J., 1949, Les coraux de l'Éocène de Bijnice-les-Bains près de Prievidza dans les Karpates Slovaques: Práce státneho geologického ústavu, 24, 3-30.
Alloiteau, J., 1952, Madréporaires post-paléozoïques, in Piveteau, J. (ed.), Traité de Paléontologie: Paris, Masson, 539-684.
Alloiteau, J., 1957, Contribution à la systématique des Madréporaires fossiles: Paris, Centre National de la Recherche Scientifique, 462 pp.
Alloiteau, J., 1958, Monographie des Madréporaires fossiles de Madagascar: Annales géologiques de Madagascar, 25, 1-218.
Angelis d'Ossat, G. de, 1905, Coralli del Cretacico inferiore della Catalogna: Palaeontographia Italica, 9, 169-251.
Babaev, R.G., Krasnov, E.V., 1977, Morphologic peculiarities of scleractinia in the connection of their habitats in Jurassic seas of the South-Eastern Caucasus and Gornogo Crimea: Trudy (Institut geologii i geofiziki. Sibirskoe otd. AN SSSR), 302, 137-148.
Baron-Szabo, R.C., 1998, A new coral fauna from the Campanian of Northern Spain (Torallola village, Prov. Lleida): Geologische und Paläontologische Mitteilungen, 23, 127-191.
Baron-Szabo, R.C., Fernández Mendiola, P.A., 1997, Cretaceous scleractinian corals from the Albian of Cabo de Ajo (Cantabria Province, N-Spain): Paläontologische Zeitschrift, 71(1/2), 35-50.
Baron-Szabo, R.C., González-León, C. M., 1999, Lower Cretaceous corals and stratigraphy of the Bisbee Group (Cerro de Oro and Lampazos areas), Sonora, Mexico: Cretaceous Research, 20, 465-497, https://doi.org/10.1006/cres.1999.0159
Baron-Szabo, R.C., Steuber, T., 1996, Korallen und Rudisten aus dem Apt im tertiären Flysch des Parnass-Gebirges bei Delphi-Arachowa: Berliner geowissenschaftliche Abhandlungen, (E) 18, 3-75.
Bataller, J., 1947, Sinopsis de las especies nuevas del Cretácico de España: Memorias de la Real Academia de Ciencias y Artes de Barcelona, (3), 28(12), 279-392.
Beauvais, M., 1961, Polypiers sénoniens des environs de Padern (Aude): Bulletin de la Société géologique de France, (7), 2(6), 723-727.
Boehm, G., 1897, Beitrag zur Gliederung der Kreide in den Venetianer Alpen: Zeitschrift der Deutschen Geologischen Gesellschaft, 49, 160-179.
Bourne, G.C., 1900, The Anthozoa, in Lankester, R. (ed.), Treatise on Zoology. Part 2: Porifera and Coelenterata: London, A. & C. Black, 59-79.
Dietrich, W.O., 1926, Steinkorallen des Malms und der Unterkreide im südlichen Deutsch-Ostafrika: Palaeontographica, (suppl.7), 1, 43-62.
Eguchi, M., 1951, Mesozoic hexacorals from Japan: Science Reports of the Tohoku Imperial University, (2: Geology), 24, 1-96.
Eliášová, H., 1976, Familles Placosmiliidae Alloiteau, 1952 et Misistellidae nov.fam. (Hexacorallia) des calcaires de Stramberk: Casopis pro Mineralogii a Geologii, 21(4), 337-347.
Ellis, J., Solander, D., 1786, The natural history of many curious and uncommon Zoophytes: London, B. White and son, 208 pp.
Étallon, A., 1864, Classe des Polypes, in Thurmann, J., Etallon, A., 1864, Lethea Bruntrutana ou études paléontologiques et stratigraphiques sur les terrains jurassiques supérieurs du Jura bernois et en particulier des environs de Porrentruy. Denkschriften der allgemeinen Schweizerischen Gesellschaft für die gesamten Naturwissenschaften, 20, 357-412.
Filkorn, H.F., Pantoja-Alor, J., 2015, Mexican Cretaceous coral species (Cnidaria, Anthozoa, Scleractinia) described as new by Filkorn & Pantoja-Alor, 2009, but deemed ‘unpublished’ under the International Code of Zoological Nomenclature: republication of data necessary for nomenclature: Bulletin of Zoological Nomenclature, 72(1), 93-101.
Fromentel, E. de, 1857, Description des polypiers fossiles de l'étage Néocomien: Bulletin de la société des sciences historiques et naturelles de l'Yonne, 1-78.
Fromentel, E. de, 1861, Introduction à l'étude des polypiers fossiles: Mémoires de la Société d'émulation du Doubs, (3), 5, 1-357.
Fromentel, E. de, 1867, Zoophytes, terrain crétacé (7), in d'Orbigny, A. (ed.), Paléontologie française, 8, 289-336.
Garberoglio, R.M., Löser, H., Lazo, D.G., 2021, Lower Cretaceous corals from the Agrio Formation, Neuquén Basin, west-central Argentina: Family Columastraeidae: Cretaceous Research, 124(104817), 1-19. https://doi.org/10.1016/j.cretres.2021.104 817
Geyer, O.F., 1955, Beiträge zur Korallenfauna des Stramberger Tithon: Paläontologische Zeitschrift, 29(3/4), 177-216.
González-León, C. M., Lucas, S.G., 1995, Stratigraphy and paleontology of the early Cretaceous Cerro de Oro Formation, central Sonora: Special Papers. Geological Society of America, 301, 41-47. https://doi.org/10.1130/0-8137-2301-9.41
González-León, O., Jeremiah, J., Schlagintweit, F., Bover Arnal, T., Moreno Bedmar, J.A., Mendoza Rosales, C.C., Vega, F.J., 2015, Novel contributions and errata to the work "Morphology and ontogeny of the fossil lobster Meyeria magna M'Coy, 1849 (Astacidae: Mecochiridae) from the Lower Cretaceous (Lower Aptian) of Mexico, United Kingdom and Spain": Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 276(3), 323-334. https://doi.org/10.1016/j.cretres.2012.03.001
Haime, J., 1852, Polypiers et Bryozoaires: Mémoires de la Société géologique de France, (2), 4(2), 279-289.
He, X., Xiao, J.-D., 1990, Jurassic and Cretaceous hexacorals of Ngari area, in Zunyi, Y., Zetong, N. (eds.), Paleontology of Ngari, Tibet (Xizang): Beijing, China University Geoscience Press, 146-159.
Hoeksema , B.W., 1989, Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae): Zoologische verhandelingen, 254, 1-295.
International Commission on Zoological Nomenclature, 1999, International code of zoological nomenclature, http://www.iczn.org/iczn/index.jsp, acceesed May 2022
Kuzmicheva, E.I., 2002, Skeletal morphology, systematics and evolution of the Scleractinia: Trudy Paleontologicheskogo Instituta, 286, 1-211.
Lamarck, J.B.P. de, 1801, Systême des animaux sans vertèbres: Paris, Comptoir géologique, 432 pp.
Lamouroux, J.V.F., 1821, Exposition méthodique des genres de l'ordre des polypiers: Paris, Agasse, 115 pp.
Liao, W.-H., Xia, J.-B., 1985, Upper Jurassic and Lower Cretaceous Scleractinia from Bangoin district of northern Xizang (Tibet): Memoirs of the Nanjing Institute of Geology and Palaeontology, 21, 119-174.
Löser, H., 2004, PaleoTax - a database program for palaeontological data: Computer & Geosciences, 30(5), 513-521, https://doi.org/10.1016/j.cageo.2004.03.009
Löser, H., 2011, The Cretaceous corals from the Bisbee Group (Sonora; Late Barremian - Early Albian): introduction and family Aulastraeoporidae: Revista Mexicana de Ciencias Geológicas, 28(2), 254-261.
Löser, H., 2013a, An Early Albian shallow marine coral fauna from Southern France – insight into evolution and palaeobiogeography of Cretaceous corals: Palaeobiodiversity and Palaeoenvironments, 93(1), 1-43, https://doi.org/10.1007/s12549-012-0088-2
Löser, H., 2013b, Revision of the Hauterivian (Early Cretaceous) corals of the Paris Basin, France: a work in progress: Bulletin d'information des géologues du Bassin de Paris, 50(1), 17-24.
Löser, H., 2016, Catalogue of Cretaceous Corals, Systematic part: Dresden, Germany, vol. 4, CPress Verlag, 710 pp.
Löser, H., 2021, Corals from the Early Cretaceous (?Late Valanginian - Aptian) of Puebla (Mexico): Family Solenocoeniidae: Paleontología Mexicana, 10(1), 37-51.
Löser, H., García-Barrera, P., Mendoza Rosales, C.C., Ortega-Hernández, J., 2013, Corals from the Early Cretaceous (Barremian - Early Albian) of Puebla (Mexico) - Introduction and family Stylinidae: Revista Mexicana de Ciencias Geológicas, 30(2), 385-403.
Löser, H., Heinrich, M., Schuster, U., 2019, Korallen von Rußbach und Gosau (Coniac-Santon; Österreich): Dresden, Germany, CPress Verlag, 367 pp.
Löser, H., Steuber, T., Löser, C., 2018, Early Cenomanian coral faunas from Nea Nikopoli (Kozani, Greece; Cretaceous): Carnets de Géologie / Notebooks on Geology, 18(3), 23-121. https://doi.org/10.4267/2042/66094
Michelin, H., 1841, Iconographie zoophytologique. Description par localités et terrains des polypiers fossiles de France (1): Paris, France, Bertrand, 40 pp.
Milaschewitsch, C., 1876, Die Korallen der Nattheimer Schichten (2): Palaeontographica, 21, 62-123.
Milne Edwards, H., Haime, J., 1848a, Observations sur les polypiers de la famille des astréides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 27(19), 465-469.
Milne Edwards, H., Haime, J., 1848b, Recherches sur les polypiers (4:) Monographie des Astréides (1:) Eusmiliens: Annales de Sciences naturelles, (3), 10, 209-320.
Milne Edwards, H., Haime, J., 1849, Mémoire sur les polypiers appartenant à la famille des oculinides, au groupe intermédiaire des Pseudoastréides et à la famille des Fongides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 29(4), 67-73.
Milne Edwards, H., Haime, J., 1851, Introduction, Corals from the Tertiary and Cretaceous, Formations, in Milne E., H., Haime, J., (ed), A monograph of the British fossil corals: London, Palaeontographical Society Monographs, 3, i-lxxxv, 1-71.
Morycowa, E., 1964, Hexacoralla des couches de Grodziszcze (Néocomien Carpathes): Acta Palaeontologica Polonica, 9(1): 1-114.
Morycowa, E., 1971, Hexacorallia et Octocorallia du Crétacé inférieur de Rarau (Carpathes orientales roumaines): Acta Palaeontologica Polonica, 16(1/2), 1-149.
Morycowa, E., Marcopoulou-Diacantoni, A., 2002, Albian corals from the Subpelagonian zone of Central Greece (Agrostylia, Parnassos region): Annales Societatis Geologorum Poloniae, 72, 1-65.
Morycowa, E., Masse, J.P., 1998, Les Scléractiniaires du Barrémien-Aptien inférieur de Provence (SE de la France): Geobios, 31(6), 725-766.
Orbigny, A. d’, 1849, Note sur les polypiers fossiles: Paris, France, Masson, 12 pp.
Prever, P.L., 1909a, Anthozoa: Memorie descrittive della carta geologica d'Italia, 5(1): 51-147.
Prever, P.L., 1909b, Coralli giurassici del Gran Sasso d'Italia: Atti della Reale Accademia di Scienze di Torino, 44, 986-1000.
Quenstedt, F.A., 1880, Petrefactenkunde Deutschlands (6:) Röhren- und Sternkorallen (2): Leipzig, Germany, Fues Verlag, 625-912.
Reig Oriol, J.M., 1991, Fauna coralina cretácica del nordeste de España: Barcelona, Spain, privately published, 53 pp.
Roniewicz, E., 1996, The key of skeletal microstructure on recognizing high-rank scleractinian taxa in the stratigraphical record: Paleontological Society Papers, 1, 187-206.
Roniewicz, E., Morycowa, E., 1993, Evolution of the Scleractinia in the light of microstructural data: Courier Forschungsinstitut Senckenberg, 164, 233-240.
Toula, F., 1889, Geologische Untersuchungen im centralen Balkan: Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Physikalische Klasse, 55, 1-108.
Turnšek, D., LeMone, D.V., Scott, R.W., 2003, Tethyan Albian corals, Cerro de Cristo Rey uplift, Chihuahua and New Mexico, in Scott, R.W. (ed.), Bob F. Perkins Memorial Volume: Special Publications in Geology, 147-185.
Vaughan, T.W., Wells, J.W., 1943, Revision of the suborders, families and genera of Scleractinia: Special Papers, Geological Society of America, 44, 1-363.
Velázquez-Heras, J.E., Castillo-Gámez, R.A., Monreal, R., Cuen-Romero, F.J., Enríquez-Ocaña, L.F., Buitrón-Sánchez, B.E., 2021, Paleoecology of a Lower Cretaceous (lower Albian) marine community of Cerro de Oro, central Sonora, Mexico: Journal of South American Earth Sciences, 113(103667), 1-11.
Wang, Y.-S., Sun, J.-B., Wang, Y., Zheng, C.-Z., Yue, Z.-Y., Liao, W.-H., 2020, Some early Cretaceous scleractinian corals from Xenkyer, Baingoin, Xizang (Tibet): Acta Palaeontologica Sinica, 59(4), 452-466.
Wells, J.W., 1932, Corals of the Trinity Group of the Commanchean of central Texas: Journal of Paleontology, 6(3), 225-256.
Wells, J.W., 1933, Corals of the Cretaceous of the Atlantic and Gulf Coastal Plains and Western Interior of the United States: Bulletins of American Paleontology, 18(67), 83-292.
Wells, J.W., 1946, Some Jurassic and Cretaceous corals from Northern Mexico: Journal of Paleontology, 20(1), 1-7.