Cómo citar: Flores-Barragan, M.A., Velasco-de León, M.P., Moreno-Bedmar, J.A., 2023, Avances en el conocimiento de la macroflora de la Formación Tuzancoa, Hidalgo, México, Pérmico inferior: Revista Mexicana de Ciencias Geológicas, v. 40, núm. 1, p. 1-15.
REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS
v. 40, núm. 1, 2023, p. 1-15
DOI: http://dx.doi.org/10.22201/cgeo.20072902e.2023.1.1694
Avances en el conocimiento de la macroflora de la Formación Tuzancoa, Hidalgo, México, Pérmico inferior
Miguel Angel Flores-Barragan1,2*, María Patricia Velasco-de León2 y Josep Anton Moreno-Bedmar3
1 Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México, Ciudad de México, México. C.P. 04510, Mexico.
2 Colección de Paleontología, Facultad de Estudios Superiores, Zaragoza, Universidad Nacional Autónoma de México, Avenida Guelatao, Ejército de Oriente, Alcaldía Iztapalapa, C.P. 09230, Ciudad de México, México.
3 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacán, C.P. 04510, Ciudad de México, México.
* 08.mike.angel@gmail.com
La paleoflora de la Formación Tuzancoa ha sido estudiada desde mediados del siglo pasado. Sin embargo, el constante descubrimiento de nuevos elementos florísticos y/o localidades hace necesaria una reevaluación del conocimiento actual de la paleoflora de esta formación. El objetivo de este trabajo es dar a conocer los nuevos reportes paleoflorísticos de tres localidades de la Formación Tuzancoa, así como analizar su distribución geográfica, cronológica y su relación con otras floras coetáneas de México. Las localidades aquí estudiadas se denominan Calnali 2, La Virgen y San Mateo 2, siendo esta última una nueva localidad. En la primera localidad se identificaron seis órdenes (Equisetales, Glossopteridales, Lepidodendrales, Marattiales, Peltaspermales y Voltziales). Mientras que en La Virgen y San Mateo 2 se identificaron cuatro órdenes (Equisetales, Lepidodendrales, Marattiales, y Peltaspermales). Los nuevos reportes corresponden a una familia, dos afinidades, cinco géneros y siete especies; de este conjunto, destacan cinco taxones que son nuevos reportes para la República Mexicana y/o importantes taxonómicamente y estos corresponden a: Majonicaceae, Odontopteris cf. brardii, Mariopteris, Taeniopteris feddeni y T. tenuis. Majonicaceae es representada por una estructura reproductora femenina probablemente relacionada con la especie Calnalia hidalguensis; Odontopteris cf. brardii es un helecho que se caracteriza por la ausencia de una vena media en sus pínnulas; Mariopteris es un helecho con pínnulas con distintos grados de fusión; Taeniopteris feddeni corresponde a una hoja entera de gran tamaño, con más de cinco cm de ancho, con una densidad de 20 a 22 venas secundarias por cm; y por último, Taeniopteris tenuis es un hoja entera con menos de cinco cm de ancho que presenta un arreglo único en su venación secundaria, con una densidad de 28 a 30 venas por cm. Estos nuevos hallazgos aumentan sustancialmente la diversidad florística de la formación y permiten establecer, junto con la litología, un ambiente transicional en Calnali 2 y un ambiente continental en La Virgen y San Mateo 2. Los rangos estratigráficos y de distribución de los taxones sugieren una edad de Pensilvánico–Pérmico inferior para los estratos portadores de plantas, además se infiere una posible afinidad geográfica con el reino Euroamericano. Por último, al comparar el listado florístico de la Formación Tuzancoa con otras unidades coetáneas con reportes de flora fósil como son las formaciones Matzitzi (Mt), Paso Hondo (Ph), Olinalá (Ol) y el Grupo Patlanoaya (Pt) se tiene que, en estas últimas (Ph, Ol y Pt) se observa una similitud mínima debido al pobre registro en relación con la Formación Tuzancoa. Con respecto a la Formación Matzitzi, las plantas indican características propias de ambientes diferentes: continental para Matzitzi y de transición-continental para Tuzancoa. Todos estos nuevos aportes nos ayudan a comprender mejor los ecosistemas terrestres de finales del Paleozoico en el margen ecuatorial occidental de Euroamérica.
Palabras clave: Pensilvánico; Pérmico; Euroamérica; taxonomía; Taeniopteris; Mariopteris; Formación Tuzancoa; Hidalgo; México.
ABSTRACT
The paleoflora of the Tuzancoa Formation has been studied since the middle of the last century. However, the constant discovery of new floristic elements and/or localities makes it necessary to re-evaluate the current knowledge of the paleoflora of this formation. Therefore, the objective of this work is to present new paleofloristic reports of three localities of the Tuzancoa Formation, as well as to analyze their geographical and chronological distribution and their relationship with other contemporary floras of Mexico. The localities studied are named Calnali 2, La Virgen, and San Mateo 2, the latter being a new locality. In the first locality, six orders were identified (Equisetales, Glossopteridales, Lepidodendrales, Marattiales, Peltaspermales and Voltziales), while in The Virgin and San Mateo 2, four orders were identified (Equisetales, Lepidodendrales, Marattiales, and Peltaspermales). The new reports correspond to one family, two affinities, five genera, and seven species; of this set, five taxa stand out as new reports for the Mexican Republic and/or as taxonomically important, and these correspond to Majonicaceae, Odontopteris cf. Brardii, Mariopteris, Taeniopteris feddeni and T. tenuis. Majonicaceae is represented by a female reproductive structure probably related to the species Calnalia hidalguensis; Odontopteris cf. Brardii is a fern characterized by the absence of a middle vein in its pinnules; Mariopteris is a fern with pinnules with varying degrees of fusion; Taeniopteris feddeni corresponds to a large entire leaf, more than five cm wide with a density of 20 to 22 secondary veins per cm; finally, Taeniopteris tenuis is an entire leaf less than 5 cm wide that presents a unique arrangement in its secondary venation with a density of 28 to 30 veins per cm. These new findings substantially increase the floristic diversity of the formation and allow to establish, together with lithology, a transitional environment in Calnali 2 and a continental environment in La Virgen and San Mateo 2. The stratigraphic and distribution ranges of the taxa, suggest an age for the plant-carrying strata of the Pennsylvanian–lower Permian; in addition, an inferred geographical affinity with the Euromerican kingdom is considered. Finally, when comparing the floristic list of the Tuzancoa Formation with other contemporary units with reports of fossil flora such as Matzitzi (Mt), Paso Hondo (Ph), Olinalá (Ol) and Patlanoaya (Pt), a minimal similarity is observed for the latter (Po, Ol and Pt) due to the poor records in relation to the Tuzancoa Fm. Regarding the Matzitzi Formation, in this case the plants indicate characteristics of different environments: Continental and humid for Matzitzi, and transitional in the locality of Calnali of the Tezuncoa Formation, while in the locality of the Virgin and San Mateo the environment is sub-humid. All these new contributions help us to better understand the terrestrial ecosystems of the late Paleozoic in the western equatorial margin of Euromerica.
Keywords: Pennsylvanian; Permian; Euramerica; taxonomy; Taeniopteris; Mariopteris; Tuzancoa Formation; Hidalgo; Mexico.
Manuscrito recibido: mayo 3, 2022
Manuscrito corregido recibido: noviembre 8, 2022
Manuscrito aceptado: noviembre 9, 2022
INTRODUCCIÓN
México tiene un extenso registro paleoflorístico que se extiende desde el Paleozoico superior hasta el Cenozoico (e.g. Silva-Pineda, 1970a, 1970b), 1984, 1987; Weber, 1997; Velasco-de León y Ortiz-Martínez, 2010. La flora paleozoica de México está representada con diferente abundancia, calidad de preservación y diversidad en ocho unidades litoestratigráficas (Silva-Pineda y Buitrón-Sánchez, 1998; Silva-Pineda et al., 2003). Para las formaciones Grupera, Santa Rosa Superior y Paso Hondo (Chiapas) solo se menciona la presencia de restos de plantas sin que se puntualice la taxonomía de algún ejemplar (Hernández-García, 1973; Silva-Pineda y Buitrón-Sánchez, 1998); en el caso de la Formación Paso Hondo, en la ilustración 31 del trabajo de Hernández-García (1973) se puede dilucidar la presencia del género Asterotheca Presl y Corda. Con respecto a la Formación Ixtaltepec del estado de Oaxaca únicamente se ha reportado una posible semilla de Pteridosperma sin tener mayores datos de esta estructura a nivel de género y/o especie. Por otro lado, para la Formación Matzitzi (Puebla), los fósiles de plantas son abundantes y presentan un buen estado de preservación registrando hasta el momento un total de 29 géneros y tres afinidades (Flores-Barragan y Velasco-de León, 2021); para el Grupo Patlanoaya (Puebla) Villaseñor-Martínez et al. (1987) identifican cinco géneros de plantas. En el caso de la Formación Olinalá (Guerrero) se han reportado tres géneros, una especie, así como una afinidad (Silva-Pineda et al., 2003). Por último, en la Formación Tuzancoa (Hidalgo) que es motivo de este estudio, los trabajos han sido intermitentes y en la mayoría de los casos se trata de material escaso y mal preservado (Silva-Pineda, 1987; Hernández-Castillo et al., 2014; Ortiz-Martínez y Velasco-de León, 2019). Como se puede observar, hasta el momento las unidades con mayor diversidad de flora fósil son las pertenecientes a las formaciones Matzitzi y Tuzancoa, esto debido en parte a la extensión de sus afloramientos y a sus ambientes de depósito que favorecieron un abundante registro de plantas (Weber, 1997; Rosales-Lagarde, 2005; Centeno-García et al., 2009); sin embargo, el descubrimiento de nuevas localidades en la Formación Tuzancoa hace necesaria una descripción del nuevo material.
La Formación Tuzancoa fue descrita anteriormente como perteneciente a la Formación Guacamaya (Carrillo-Bravo, 1965), para posteriormente ser propuesta como una unidad independiente (Rosales-Lagarde et al., 2005). Esta formación se describe como una secuencia siliciclástica con brechas andesíticas intercaladas con arenisca y lutita de origen volcánico, caracterizada por su registro fósil de bivalvos, crinoideos y foraminíferos. Todos estos datos indican un ambiente de depósito marino somero y transicional (Carrillo-Bravo, 1965; Rosales-Lagarde et al., 2005; Sour-Tovar et al., 2005; Quiroz-Barroso et al., 2012; Buitrón-Sánchez et al., 2017; Gutiérrez-Alejandre, 2018). Desde el punto de vista paleobotánico se tienen como antecedentes algunos trabajos taxonómicos (Silva-Pineda, 1963; 1987; Hernández-Castillo et al., 2014; Ortiz-Martínez y Velasco-de León, 2019; Velasco-de León et al., 2020). La sección tipo de esta formación aflora en las cercanías del poblado Otlamalacatla, Hidalgo (Rosales-Lagarde et al., 2005). En esta área y en la zona que comprende los poblados Calnali-Tlahuelompa se han realizado diversos estudios taxonómicos, cronológicos y biogeográficos sobre la fauna marina (Sour-Tovar et al., 2005; Quiroz-Barroso et al., 2012; Buitrón-Sánchez et al, 2017; Gutiérrez-Alejandre, 2018). El alcance estratigráfico de las especies de bivalvos, braquiópodos, gasterópodos y trilobites descritas para esta unidad han permitido asignar una edad de Pérmico temprano (Cisuraliense) y proponer una afinidad de esta fauna con la provincia Grandian de Norteamérica (Quiroz-Barroso et al., 2012).
El primer estudio paleobotánico para esta formación se remonta a 1963 (Silva-Pineda, 1963); estudios posteriores dieron cuenta de que este análisis taxonómico comprendía material de distintas localidades que difícilmente pertenecen únicamente a la Formación Tuzancoa. Los resultados de este primer trabajo fueron reevaluados por Silva-Pineda (1987), en el que se registraron cinco géneros y una afinidad cf. Gangamopteris McCoy; Neuropteris Brongniart (Sternberg); Odontopteris Brongniart; Pecopteris Brongniart; Taeniopteris Brongniart y Walchia Sternberg pertenecientes a una localidad cercana al poblado de Calnali, Hidalgo (Calnali 1). Posteriormente Weber (1997) incrementa los reportes para la Formación Tuzancoa con los registros de Sigillaria Brongniart y Cyperites Lindley y Hutton pertenecientes a una nueva localidad cercana al poblado de San Mateo, Hidalgo (San Mateo 1) y propuso cierta similitud con la flora de la Formación Matzitzi.
A inicios de este siglo en la localidad Calnali 1 se describió un nuevo género y especie de conífera, Calnalia hidalguensis Hernández-Castillo, Silva-Pineda y Cevallos-Ferriz (Hernández-Castillo et al., 2014), y por su posición con respecto a estratos portadores de invertebrados se le asignó a esta especie una edad de Pérmico temprano. Posteriormente se reportaron dos nuevas localidades; la primera de ellas, denominada Calnali 2, aflora en los alrededores del poblado del mismo nombre y la segunda llamada La Virgen cercana al poblado de Jalapa, Hidalgo (Figura 1). En la primera de ellas se describen semillas de los géneros Cordaicarpus Geinitz y Cornucarpus Arber (Ortiz-Martínez y Velasco-de León, 2019), mientras que en la segunda se dan a conocer datos preliminares de la paleoflora, describiendo un total de nueve géneros, una afinidad, un fosilitaxón, además de una especie (Velasco-de León et al., 2020). Sin embargo, en estas nuevas localidades la ausencia de estudios taxonómicos de invertebrados hace difícil estimar la edad de las rocas, por lo que un primer acercamiento para subsanar esta problemática se basa en el análisis del alcance estratigráfico de algunos taxones de plantas.
Por lo mencionado previamente, el objetivo de este trabajo es actualizar el listado florístico de la Formación Tuzancoa, correspondiente a tres localidades. Además, se analizan las implicaciones cronoestratigráficas y paleobiogeográficas de la flora estudiada y su correlación con la paleoflora de otras unidades del Paleozoico superior de México.
ZONA DE ESTUDIO
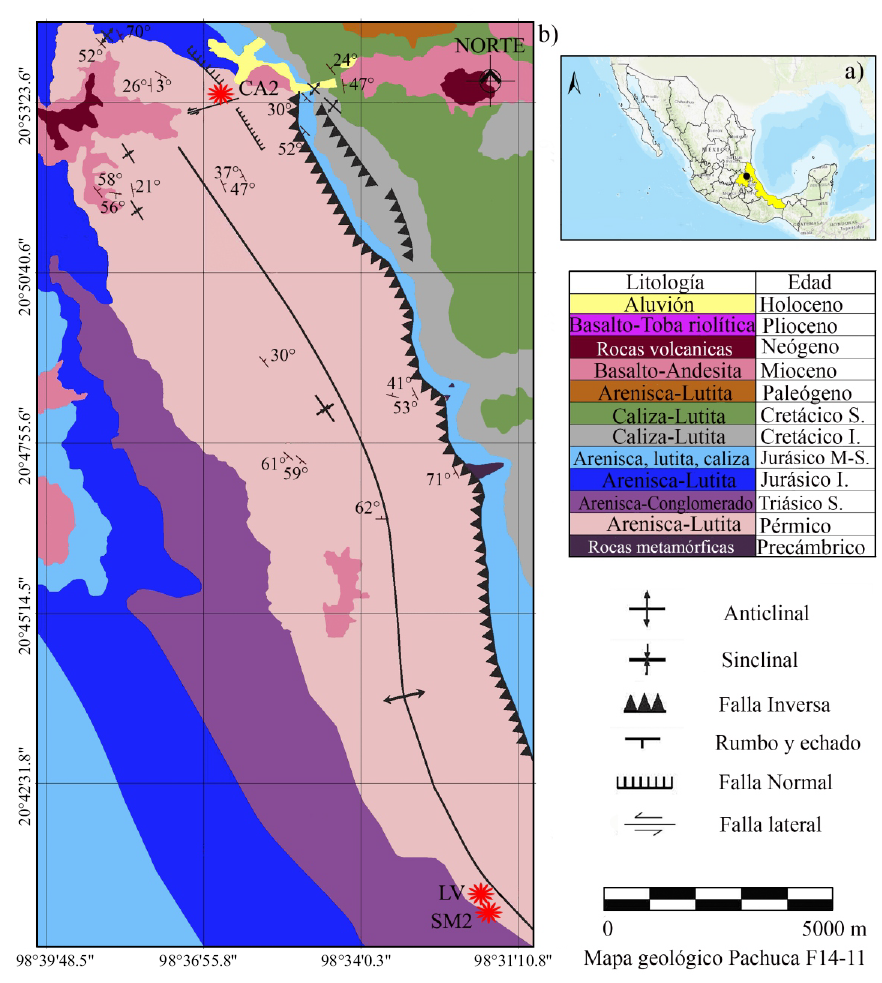
El material aquí revisado pertenece a tres localidades de la Formación Tuzancoa que se ubican en el sector centro-sur del anticlinorio de Huayacocotla en los estados de Hidalgo y Veracruz. La primera localidad corresponde a Calnali 2 y se ubica aproximadamente 3 km al oeste de la ciudad del mismo nombre (20°53'36.3"N 98°36'34.4"W). La segunda localidad nombrada La Virgen se ubica 1 km al oeste del poblado de Jalapa a 13.9 km al noroeste de Zacualtipán (20°40'35''N, 98°31'34.6''W). Cabe mencionar que estas dos localidades se encuentran en el estado de Hidalgo. Por último, se da a conocer la nueva localidad nombrada San Mateo 2 localizada a 1 kilometro al norte del poblado del mismo nombre, perteneciente al estado de Veracruz (20°40'35''N, 98°32'3.2''W) (Figura 1).
Figura 1. Mapa de ubicación de las zonas de estudio. a) Mapa de la República Mexicana mostrando en amarillo los estados de Hidalgo y Veracruz donde se ubican las zonas de estudio representadas con el punto negro. b) Mapa geológico donde se ubican las tres localidades estudiadas en este trabajo, CA2: Calnali 2; LV: La Virgen; SM2: San Mateo 2. Simbología: I) Inferior. M) Medio y S) Superior. Tomado y modificado de Velasco-de León et al. (2020).
MATERIAL Y MÉTODO
Se realizaron tres campañas de campo en las localidades Calnali 2 (Ca2); La Virgen (LV) y San Mateo 2 (SM2) pertenecientes a la Formación Tuzancoa donde se recolectaron en total 402 ejemplares; los fósiles corresponden a impresiones en lutita y arenisca de grano fino a mediano. Se levantaron cuatro secciones estratigráficas para ubicar los estratos portadores de plantas. Con una vara de Jacob se midieron los espesores de los estratos a la vez que se puso especial atención a los cambios litológicos y granulométricos, así como a estructuras sedimentarias primarias. El material fósil colectado esta resguardado en la Colección de Paleontología de la Facultad de Estudios Superiores de Zaragoza (CFZ-Tz). Con la finalidad de obtener mayor precisión en la medición de los caracteres morfológicos empleados en la identificación se usó el programa Image J. La asignación taxonómica del material aquí analizado se basó en trabajos que emplearon caracteres morfológicos de las láminas o figurados en Remy y Remy (1975), Artabe (1985) y van Konijnenburg-van Cittert et al. (2017).
RESULTADOS Y DISCUSIÓN
Litología
En la localidad CA2 se midió una sección estratigráfica de 123.4 m. En los primeros 21 metros aflora una secuencia donde se intercalan lodolita y limolita; en esta zona se ubican estratos portadores de plantas e invertebrados. Los suprayacen una serie de estratos de 31 metros de arenisca de grano fino-mediano con laminación paralela. Los siguientes 23 m corresponden a una secuencia alterna de arcilla y limolita, en el metro 111 es donde se ubica la última zona de recolecta de esta localidad (Figura 2). A lo largo de esta columna se observan cambios en la energía de transporte de los sedimentos que se manifiestan mediante cambios granulométricos que se pueden observar y algunas estructuras sedimentarias como es la laminación cruzada.
Para esta área de estudio existen trabajos geológicos que infieren un ambiente de depósito relacionado con ascenso y descenso del nivel del mar, que generaron transgresiones y regresiones marinas en las llanuras costeras del continente (Gutiérrez-Alejandre, 2018; Velasco-de León et al., 2020).
Con respecto a la localidad LV (Figura 3a-3b) se observa una secuencia de rocas de casi tres metros de espesor, a lo largo de la cual se ubican diferentes zonas con material fósil. En la base se observa una alternancia de lutita, limolita y arenisca de grano fino en estratos de 20 cm de espesor con laminación paralela; arriba de esta arenisca laminada se ubica un paquete de estratos delgados de arenisca de grano mediano con un espesor de 40 cm. Lo suprayace un paquete de rocas de 1 m de espesor, que en su base tiene capas onduladas de 3 a 7 cm de espesor con una gradación inversa de arenisca de grano fino-mediano. Posteriormente se ubica una arenisca de grano fino a mediano con laminación paralela. La cima de la sección estratigráfica mide unos 60 cm de espesor con arenisca muy fina-fina, con abundante flora fósil y con presencia de laminación tipo flaser (Figura 3b). Para esta localidad se ha propuesta un ambiente de depósito meramente continental a partir de las estructuras sedimentarias y su contenido fósil que es claramente autóctono (Velasco-de León et al., 2020).
Por último, en la localidad SM2 (Figura 3c-3f), se midieron dos secciones estratigráficas debido a que una falla no permite observar la secuencia sedimentaria completa. La sección 1, tiene un espesor 6.50 m, con una granulometría que puede ir de limo-arena de grano muy fino a arenisca de grano muy fino-mediano con presencia de laminación paralela tenue; la zona de recolecta se ubica entre los metros tres y cuatro de la sección estratigráfica (Figura 3d). La sección número 2 (Figura 3e-3f), tiene un espesor de apenas 2.50 m; en ésta se observa una intercalación de limolita con estratos más delgados de arenisca de grano fino, y en la cima de la sección se observa laminación paralela; se ubicaron dos zonas de recolecta, pero dicho material es completamente distinto al recolectado en la sección número 1. A partir de las intercalaciones de limo con arena, así como la laminación interna de ambas secciones se pueden asignar como parte de la facies Fl que se interpretan como canales abandonados o depósitos de inundación menguantes (Miall, 2006).

Figura 2. Sección estratigráfica de Calnali 2, Formación Tuzancoa. a) sección estratigráfica. b y c) Fotos de la zona de recolecta del material fósil, donde se observan la secuencia alterna de lodolita y limolita. Simbología: 1) Arcilla. 2) Limo. 3) Muy Fino. 4) Fino. 5) Mediano. 6) Grueso y 7) Muy Grueso. Escala 1 metro.

Figura 3. Secciones estratigráficas de las localidades La Virgen y San Mateo 2. a) Localidad La Virgen. b) Foto del afloramiento donde se observa en la base una alternancia de lutita, limolita y arenisca y en la parte media y superior los estratos de arenisca. c) Localidad San Mateo 2, sección 1. d) Foto de los estratos portadores de plantas y la secuencia de limolita-arenisca. e) Localidad San Mateo 2, sección 2. Simbología: 1) Arcilla. 2) Limo. 3) Muy Fino. 4) Fino. 5) Mediano. 6) Grueso y 7) Muy Grueso. Escala 1 metro.
Paleoflora
En la localidad Ca2 se identificaron un total de 132 ejemplares que fueron ubicados en seis órdenes, 11 géneros y seis especies, siendo Calnalia Hernández-Castillo, Silva-Pineda y Cevallos Ferriz y Asterotheca (Figura 4c) los géneros más abundantes. En la localidad LV se identificaron 194 ejemplares asignados a 4 órdenes, 10 géneros y seis especies; en esta área destaca la presencia de Asolanus Wood (Figura 4a) Comia Zalessky (Figura 4b), Mariopteris Zeiller, y Taeniopteris. Por último, en la localidad SM2 se identificaron 76 fósiles que se corresponde a 4 órdenes 13 géneros y siete especies; Calamites Suckow y Taeniopteris son los géneros más abundantes (Tabla 1).
A continuación, se describen los nuevos registros para estas localidades.

Figura 4. Macroflora del Paleozoico superior de México, Formación Tuzancoa. a) Corteza de Asolanus; la flecha indica la zona de inserción de las hojas. b) Hoja de Comia, la flecha indica la venación característica de este género. c) Pinna del género Asterotheca. e) Pínnula de la especie Odontopteris cf. O. brardii. d) Dibujo de la imagen e, donde se observa la venación típica de este género. f) Pinna incompleta de Odontopteris cf. O. brardii. g) Estructura reproductora de la familia Majonicaceae. h) Dibujos donde se observa el número variable de brácteas presentes en la famila Majonicaceae. j) Estructura femenina, donde la flecha indica la longitud de las bracteas. Simbología: 1) Raquis. 2) Margen. 3) Bracteas. Escala 1 cm.
PALEONTOLOGÍA SISTEMÁTICA
Orden: Voltziales Stewart et Rothwell, 1993
Familia: Majonicaceae Clement-Westerhof 1987 (Figura 4g-4j)
Ejemplares. CFZ Tuz-51, 63, 86, 122.
Órgano. Estructura reproductora femenina.
Ocurrencia. Localidad Calnali 2 (Hidalgo).
Descripción. Brote enano, el más grande con 1 cm de largo por 1.3 cm
de ancho, aplanados y simétricos bilateralmente, con un máximo de ocho brácteas fusionadas en su base; cada bráctea mide 0.8 cm de largo por 0.1 cm de ancho, estriadas longitudinalmente que terminan en un ápice agudo, brácteas sin presentar conexión con esporofilos.
Observaciones. Dentro del orden de las Voltziales se encuentra la familia Majonicaceae que incluye estructuras reproductoras femeninas, agrupadas en un brote enano formado por escamas estériles individuales más o menos libres en número variable y escamas ovulíferas (esporofilos) (Forte et al., 2017). Sin embargo, en este caso al no poder observar escamas fértiles y por consiguiente las áreas de unión de los óvulos, así como su arreglo con respecto a las escamas estériles, se hace imposible la identificación en un rango taxonómico inferior por lo cual únicamente se asigna a nivel de familia. Este es un elemento típico del Paleozoico superior de Euroamérica, sobre todo en su parte ecuatorial (Looy, 2007).
Orden Peltaspermales Taylor, 1981
Género Odontopteris (Brongniart) Sternberg, 1825
Odontopteris cf. brardii Brongniart, 1822 (Figura 4d-4f)
Ejemplares. CFZ Tuz-55, 59, 97, 309, 315, 326, 364, 376, 398, 403, 404.
Órgano. Pinna incompleta.
Ocurrencia. Localidad Calnali 2 (Hidalgo), La Virgen (Hidalgo) y San Mateo 2 (Veracruz).
Descripción. Pinna incompleta, la más completa con 2.2 cm de largo por 1.9 cm de ancho; pínnulas de 0.7 cm de largo por 0.5 cm de ancho, con un arreglo de opuestas a sub-opuestas, triangulares, ligeramente falcadas, unidas por toda su base al raquis, muy cercanas entre sí, sin fusionarse, de margen entero con ápice obtuso; las pínnulas no presentan vena media y se observa una densidad de venación en el margen de 25 venas por centímetro, divididas dicotómicamente.
Observaciones. Odontopteris es un género creado para agrupar pínnulas que están totalmente adheridas al eje y carecen de una vena media, en su lugar están vascularizadas por varias venas que se insertan desde la base y se pueden bifurcar a intervalos irregulares en su curso hacia el margen (Knight, 1983). Debido a la forma general de las pínnulas, así como su tamaño y venación tiene similitud a la especie O. brardii, sin embargo, debido a que se carece de ejemplares más completos no se puede asegurar dicha asignación, por tal motivo se maneja únicamente como afinidad. Esta especie está ampliamente distribuida en Euroamérica durante todo el Carbonífero (Cleal y Shute 1991).
Orden Incertae sedis
Familia Mariopteridaceae Němejc 1968
Género Mariopteris Zeiller 1879
Mariopteris sp. (Figura 5a-5d)
Ejemplares. CFZ Tuz-8, 13, 17, 25, 32 (1) (2), 36, 37, 113, 142, 144, 151, 158, 163, 170, 172, 174 (1) (2), 177, 180, 182, 183, 185, 190-196, 198-207, 209, 210, 213-218, 220, 221, 223-225, 228-230, 232, 235, 237, 240, 241, 242-247, 249-252, 259, 260, 267, 272, 276, 278, 279, 282, 285, 289, 292, 296-300, 302, 304, 305, 390, 411-414 y 416-418.
Órgano. Frondas completas.
Ocurrencia. Localidad La Virgen (Hidalgo) y San Mateo 2 (Veracruz).
Descripción. Fronda incompleta, estéril, probablemente bipinnada-tripinnada, raquis primario de 0.7 cm de ancho recorrido longitudinalmente por varias estrías, pinnas primarias incompletas de al menos 7 cm de largo por 6.5 cm de ancho, las pinnas se insertan en un ángulo de 40°–60° con respecto al raquis, generalmente opuestas. Raquis secundario estriado longitudinalmente de 0.2 cm de oinsertan en un ángulo variable con respecto al raquis, generalmente opuestas.
Pínnulas triangulares, unidas al raquis por toda su base, presentan ápice de agudo a obtuso, pínnulas grandes de 0.5 cm de largo por 0.2 cm de ancho o más pequeñas de 0.3 cm de largo por 0.2 cm de ancho, muy juntas entre sí, fusionadas parcial o totalmente; presencia de pínnulas asimétricas en la base de las pinnas. Vena media oblicua, que no recorre más de la mitad de la pínnula, en algunos casos se presenta una segunda vena que se inserta directamente del raquis, una en la parte central de la pínnula (vena media) y otra en la parte basiscópica de la misma, la vena media se dicotomiza de 2 a 4 veces, mientras que la basiscópica únicamente se divide de 1 a 2 veces; aunque el número de divisiones varía dependiendo del tamaño y grado de fusión de las pínnulas.
Observaciones. Anteriormente algunos de estos ejemplares habían sido asignados al género Lobatopteris Wagner; sin embargo, este género se utiliza para ubicar helechos tri-quintupinnados, con una venación muy característica, debido a que cada vena secundaria se divide una vez, bifurcándose posteriormente la vena superior; formando una venación trífida. A continuación, se divide asimismo la vena intermedia originándose así, venación cuadrífida (Wagner, 1959). Este arreglo en la venación no es observable en los ejemplares aquí analizados, por lo que se descarta la presencia de este género para la Formación Tuzancoa.
La familia Mariopteridaceae se distribuyó ampliamente durante el Pensilvánico-Pérmico inferior de Euroamérica (Taylor et al., 2009) se distingue por la presencia de pínnulas asimétricas en la base de las pinnas en su porción basiscópica; además de ganchos trepadores especializados que generalmente se desarrollan a partir de prolongaciones apicales (Gastaldo y Boersma, 1983). Esta familia se puede dividir principalmente en tres géneros, Karinopteris Boersma (Namuriano-Westfaliano), Mariopteris (Namuriano-Westfaliano) y Pseudomariopteris Danze y Corsin (Stephaniano-Cisuraliano) (Taylor et al., 2009).
En general los tres géneros se diferencian en la segmentación de las frondas, cuadripartitas en Mariopteris y bipartitas en Karinopteris y Pseudomariopteris. Sin embargo, aunque se tiene gran número de ejemplares en la Formación Tuzancoa, ninguno de ellos permite observar el arreglo de segmentación de la fronda. Con respecto a las pinnas en la mayoría de las especies de Karinopteris y Mariopteris, la porción proximal de la hoja es tripinnada y la porción distal bipinnada, mientras que en Pseudomariopteris toda la fronda es bipinnada (Taylor et al., 2009). En los ejemplares aquí analizados se observa en algunos casos como la fronda pasa de tripinnada a bipinnada, debido a la completa fusión de las pínulas, por lo cual se descarta que pertenezca al género Pseudomariopteris.
Por otro lado, Karinopteris se caracteriza por presentar pínnulas esfenopteroides, constreñidas en la base, que si bien presentan una variable forma de lobulación esta nunca es redonda. Mientras que en Mariopteris se presentan pínnulas libres, parcial o completamente fusionadas que se insertan por toda su base; la forma de las pínnulas es ancha o estrechamente triangular a ovada, con ápices redondeados a agudos.
Los márgenes de las pínnulas pueden ser enteros, dentados o lobulados. La venación se compone de una vena media decurrente que se extiende hasta al menos la mitad de la longitud de la pínnula, las venas secundarias se bifurcan al menos una vez (Gastaldo y Boersma, 1983). Dicho lo anterior, los ejemplares de Hidalgo y Veracruz son asignables a al género Mariopteris, sin embargo, se necesitan ejemplares mejor conservados que permita observar las segmentaciones de las frondas, así como los ganchos trepadores de las pinnas, para confirmar la asignación a este género.
Orden Incertae sedis
Género Taeniopteris Brogniart 1828
Taeniopteris feddeni Zeiller 1882 (Figura 5e-5g)
Ejemplares. CFZ Tuz-155, 156, 157, 159, 175, 176, 239 (1) (2), 258.
Órgano. Hoja incompleta.
Ocurrencia. Localidad La Virgen (Hidalgo).
Descripción. Hoja entera, incompleta, con un largo máximo conservado de 8.3 cm y 8.2 cm de ancho, de margen entero, ápice en forma de U, con un fuerte raquis estriado de 0.3 cm. Las venas son perpendiculares al raquis en un ángulo cercano a 90°, presentan pocas dicotomías de manera irregular a lo largo de toda la hoja, teniendo una densidad de venación de 20 a 22 venas por centímetro.
Observaciones. En el registro fósil las hojas enteras, recorridas por un raquis, que carecen de caracteres cuticulares y reproductores se ubican generalmente en el fósilitaxón Taeniopteris (van Konijnenburg-van Cittert et al., 2017). Las observaciones indican similitud con la especie Taeniopteris feddeni que se caracteriza por presentar una lámina muy ancha, así como por el arreglo y densidad de venación de 20 a 22 por cm. Una especie similar a esta es T. abnormis Remy y Remy ya que presenta un tamaño y arreglo de las venas muy similar, sin embargo, se diferencia por una densidad mayor por centímetro en esta última (Remy y Remy, 1975); por lo cual, los ejemplares de Hidalgo se asignan a la especie T. feddeni. Este taxón se ha descrito para el Pérmico de la India (Bose et al., 1990).
Taeniopteris tenuis Doubinger y Vetter, 1959 (Figura 5h-5i)
Ejemplares. CFZ Tuz-320
Órgano. Hoja incompleta
Ocurrencia. Localidad Calnali (Hidalgo)
Descripción. Hoja entera, incompleta, con un largo máximo conservado de 3.5 cm y un ancho estimado de 3 cm, sin presencia de base y ápice, de margen entero; presenta un raquis estriado de 0.15 cm. Las venas se insertan en un ángulo de 80-90°, dicha venación presenta de dos a tres dicotomías muy cerca del raquis para posteriormente recorrer la lámina de manera paralela; la hoja presenta una densidad de venación de 28 a 30 venas por centímetro.
Observaciones: El ejemplar de Hidalgo presenta una hoja ligeramente más ancha a lo descrito en la literatura, todas las demás características concuerdan con la especie T. tenuis, por lo cual se asigna el único ejemplar a dicha especie. Otro taxón muy similar a esta especie corresponde a T. carnoti Renault y Zeiller, pero se deferencia por presentar dicotomías en la base y parte media de la hoja. T. tenuis se ha descrito para el Carbonífero superior de Europa (Doubinger y Vetter, 1959).
Tabla 1. Tabla de diversidad, abundancia, rangos de edad y de distribución de los especímenes reportados para la Formación Tuzancoa. En la columna de distribución las letras corresponden a: A) reino de Angara. C) reino de Cathaysian E) reino de Euroamérica y G) reino de Gondwana. El porcentaje de diversidad se obtuvo a partir del número de ejemplares asignados a un taxon determinado entre el total de ejemplares.
|
Género |
Especie |
Localidad |
Rango estratigráfico mínimo |
Rango estratigráfico máximo |
Distribución |
Porcentaje de diversidad (%) |
|||
|---|---|---|---|---|---|---|---|---|---|
|
Calnali 2 |
La Virgen |
San Mateo 2 |
|||||||
|
Equisetales |
Calamites |
Calamites approximatus Schlotheim |
* |
* |
Pensilvánico |
Pensilvánico |
E |
1.4 |
|
|
Equisetales |
Calamites |
Calamites cistiiformis Stur |
* |
* |
Misisípico |
Pensilvánico |
E |
2.2 |
|
|
Equisetales |
Calamites |
Calamites varians Sternberg |
* |
* |
Pensilvánico |
Pensilvánico |
E |
4.4 |
|
|
Glossopteridales |
Gangamopteris |
Gangamopteris sp. |
* |
Pérmico temprano |
Pérmico tardío |
G |
0.7 |
||
|
Insertae sedis |
Cordaicarpus |
Cordaicarpus sp. |
* |
* |
Misisípico |
Triásico tardío |
A-C-E-G |
3.2 |
|
|
Insertae sedis |
Cornucarpus |
Cornocarpus sp. |
* |
Pensilvánico |
Pérmico temprano |
A-C-E-G |
0.2 |
||
|
Insertae sedis |
Taeniopteris |
Taeniopteris cf. magnifolia Rogers |
* |
Triasico medio |
Triásico tardío |
G-E |
1.9 |
||
|
Insertae sedis |
Taeniopteris |
Taeniopteris feddeni |
* |
Pérmico temprano |
Pérmico temprano |
G |
2.2 |
||
|
Insertae sedis |
Taeniopteris |
Taeniopteris multinervia Weiss |
* |
* |
* |
Pensilvánico |
Triásico |
E |
12.6 |
|
Insertae sedis |
Taeniopteris |
Taeniopteris tenuis |
* |
Pensilvánico |
Pensilvánico |
E |
0.2 |
||
|
Lepidodendrales |
Asolanus |
Asolanus sp. |
* |
Pensilvánico |
Pérmico temprano |
E |
0.4 |
||
|
Lepidodendrales |
Cyperites |
Cyperites bicarinatus Lindley et Hutton |
* |
* |
* |
Misisípico |
Pensilvánico |
E |
2.9 |
|
Lepidodendrales |
Knorria |
Knorria sp. |
* |
* |
Misisípico |
Pérmico medio |
A-E |
0.7 |
|
|
Lepidodendrales |
Sigillaria |
Sigillaria brardii Brongniart |
* |
Pensilvánico |
Pérmico temprano |
C-E |
1.9 |
||
|
Lepidodendrales |
Stigmaria |
Stigmaria sp. |
* |
* |
Misisípico |
Pérmico tardío |
C-E |
0.9 |
|
|
Lepidodendrales |
Syringodendron |
Syringodendron sp. |
* |
Pensilvánico |
Pérmico tardío |
C-E |
0.2 |
||
|
Marattiales |
Asterotheca |
Asterotheca cyathea |
* |
* |
* |
Pensilvánico |
Pensilvánico |
E |
10.4 |
|
Marattiales |
Diplazites |
Diplazites sp. |
* |
* |
Misisípico |
Pérmico tardío |
E-C |
1.9 |
|
|
Mariopteridaceae |
Mariopteris |
Mariopteris sp. |
* |
* |
Pensilvánico |
Pérmico temprano |
C-E |
24.3 |
|
|
Peltaspermales |
cf. Macroneuropteris Cleal, Shute et Zodrow |
* |
* |
Pensilvánico |
Pensilvánico |
E |
4.4 |
||
|
Peltaspermales |
Comia |
Comia sp. |
* |
* |
* |
Pérmico temprano |
Pérmico tardío |
A-C-E |
6.9 |
|
Peltaspermales |
cf. Linopteris Presl emend. Gothan |
* |
Pensilvánico |
Pensilvánico |
E |
0.2 |
|||
|
Peltaspermales |
Neuropteris |
Neuropteris sp. |
* |
Pensilvánico |
Triásico temprano |
A-C-E |
0.9 |
||
|
Peltaspermales |
Odontopteris |
odontopteris cf. brardii |
* |
* |
Pensilvánico |
Pensilvánico |
E |
2.7 |
|
|
Voltziales |
Calnalia |
Calnalia hidalguensis |
* |
Pérmico temprano |
Pérmico temprano |
E |
9.7 |
||
|
Majonicaceae |
* |
Pérmico temprano |
Pérmico tardío |
E |
0.9 |
||||

Figura 5. a) Fronda de Mariopteris la flecha indica una pínnula decurrente. b) Acercamiento de una pínnula de Mariopteris la flecha indica su ápice imparipinnado. c) Dibujo de una pínnula de Mariopteris aun sin fusionar. d) Dibujo de una pínnula de Mariopteris fusionada. e) Dibujo que muestra el margen y la venación de Taeniopteris feddeni. f) Hoja de Taeniopteris feddeni, donde se observa la forma de su ápice. g) Hoja incompleta de Taeniopteris feddeni de gran tamaño, además de un acercamiento a la venación. h) Foto de una hoja incompleta de Taeniopteris tenuis. i) Dibujo que muestra el margen y la venación de Taeniopteris tenuis. Simbología: 1) Raquis. 2) Margen. 4) Vena media. Escala 1 cm.
ANÁLISIS TAXONÓMICO DE LA PALEOFLORA DE LA FORMACIÓN TUZANCOA
Para la Formación Tuzancoa existen hasta el momento cinco trabajos paleobotánicos; el primero de ellos con material que probablemente pertenece a dos unidades distintas: formaciones Huizachal y Guacamaya, actualmente se sabe que esta última unidad aflora únicamente en el Estado de Tamaulipas (Silva-Pineda, 1963; Rosales-Lagarde et al., 2005); posteriormente, el listado se restringe a la Formación Tuzancoa (Silva-Pineda, 1987). Este último trabajo junto con el de Hernández-Castillo et al. (2014) se centran en el estudio de localidad Calnali 1 no encontrada por los autores de este trabajo, quienes consideran que la localidad nombrada Canali 2 es correlacionable a ésta, debido a la litología y a los grupos taxonómicos dominantes de ambas. En la localidad Calnali 2, Ortiz-Martínez y Velasco-de León, (2019) identifican dos géneros de semillas descritas para el Carbonífero. En total, los trabajos realizados suman siete géneros y una afinidad para estas dos localidades. En esta investigación se aumenta en casi un 50 % el listado florístico, documentando una nueva familia, cuatro géneros, y una afinidad genérica, así como seis especies (Tabla 1). Por último, a partir de parámetros como la abundancia de los taxones, tamaño de los órganos (hojas, tallos y estructuras reproductoras) y grado de fragmentación (Greenwood, 1991), se infiere que el material de esta localidad es alóctono.
Por otra parte, Velasco-de León et al. (2020) describen la nueva localidad llamada La Virgen que se caracteriza por poseer paleoflora con un alto grado de preservación (presencia de cutícula), que junto con la abundancia y el tamaño de las hojas, frondas y cortezas encontrados se puede interpretar como una flora autóctona (Grennwood, 1991; Gastaldo, 1988). El listado original reportado para esta localidad (La Virgen) se corrige en este trabajo. Previamente se discutieron los caracteres que permiten identificar al género Mariopteris anteriormente descrito como Lobatopteris. Algo similar sucede con las ramas asignadas anteriormente a Calnalia ya que en ningún caso es posible observar hojas escuamiformes rodeando el tallo que permiten una posible determinación (Hernández-Castillo et al., 2014), por lo que se tratan únicamente de raicillas indeterminadas.
Con respecto a la localidad San Mateo 1, ésta fue estudiada por Weber (1997) quien menciona la presencia de dos géneros (Cyperites y Sigillaria). Nuevamente esta localidad es similar a San Mateo 2, debido a la coincidencia de los grupos taxonómicos, por lo cual se consideran correlacionables. Por último, a partir de los parámetros de Gastaldo, (1988) y Greenwood (1991) se considera al material autóctono.
En total, en la Formación Tuzancoa se identificaron seis ordenes, 18 géneros y 10 especies; todos estos taxones son típicos de finales del Paleozoico y adaptados a un clima generalmente húmedo con periodos de estrés hídrico (Thomas, 1978; Phillips y DiMichele, 1992; DiMichele et al., 2006; Looy, 2007; Taylor et al., 2009; DiMichele et al., 2010; Wilson et al., 2017). Sin embargo, las localidades La Virgen y San Mateo 2 que tienen material autóctono presentan mayor similitud, ya que comparten casi el 40 % de los taxa (Tabla 1). A diferencia de Calnali 2 con la que únicamente comparten el 15 % de su flora. Se propone que la diferencia entre estas floras se debe a un sesgo tafonómico más que a las condiciones ambientales en las que se desarrollaron y se consideran una misma flora.
BIOESTRATIGRAFÍA Y PALEOBIOGEOGRAFÍA DE LA FORMACIÓN TUZANCOA
Como se observa en la Tabla 1, los géneros y especies descritos para la Formación Tuzancoa son característicos del Paleozoico superior. Al observar los rangos de distribución de los distintos géneros (dejando por el momento afuera las afinidades genéricas o superiores) se observa que el mayor rango de coincidencia se da hacia el Pensilvánico ya que 19 de los 23 fosilitaxones se distribuyen durante este periodo. Sin embargo, es importante señalar que la diversidad tanto de Equisetales y Lepidodendrales (8 % y 7 % respectivamente) no es acorde a una flora típica de este periodo como se observa en otras localidades de Norteamérica (Thomas, 1978; Phillips y DiMichele, 1992; Wilson et al., 2017). En su lugar destacan taxones de una edad más joven (Pérmico) por ejemplo Calnalia, Comia y Gangamopteris, en el caso de Calnalia con una abundancia del 30 %. El segundo mayor rango de coincidencia se da hacia el Pérmico temprano (15 de 23 taxones); aunado a esto, los porcentajes de coincidencia que se muestran en la Tabla 1 permiten proponer una edad de Pensilvánico-Pérmico temprano para los estratos portadores de plantas de la Formación Tuzancoa.
Por último, esta propuesta de edad es acorde con los registros de invertebrados como lo indican Babylonites sp. Yochelson, Strobeus welleri Knight (gasterópodos) o Euchondria cooperi Newell y Boyd y Acanthopecten coloradoensis Newberry (Bivalvos) (Rosales-Lagarde et al., 2005; Sour-Tovar et al., 2005; Quiroz-Barroso et al., 2012), que se extienden desde el Pensilvánico superior hasta el Pérmico medio.
Con respecto a la distribución de la paleoflora de la Formación Tuzancoa, se observa que 11 taxones tienen una distribución cosmopolita; de estos, 11 géneros son Euroamericanos y solo en dos casos (Gangamopteris sp. y Taeniopteris feddeni) corresponden a Gondwana con casi el 3 % de representatividad. Estos datos coinciden nuevamente con los resultados obtenidos a partir del análisis de gasterópodos y bivalvos (Quiroz-Barroso et al., 2012). Propuestas de una biota con afinidad a Euroamérica han sido realizadas por otros autores (Quiroz-Barroso et al., 2000; Navarro-Santillán et al., 2002), quienes han propuesto que durante el Devónico-Pérmico, el microcontinente Oaxaquia colisionó con Laurasia, para posteriormente crearse en el este de México un arco continental que conectaba al norte con el cinturón Ouachita y al sur con la región de Centroamérica y andina que permitió la migración de animales (p. ej. gasterópodos y bivalvos) y plantas (p. ej. Gangamopteris) entre distintos reinos, sobre todo entre Euroamérica y Gondwana (Ortega-Gutiérrez et al., 1995; Vachard y Pantoja, 1997; Vachard et al., 2000; Centeno-García, 2005).
Por último, la litología de las distintas zonas de colecta aquí descritas muestran un ambiente transicional (Calnali 2) a completamente continental (La Virgen y San Mateo 2). Hasta el momento se desconoce su relación con las cinco facies marinas propuestas para esta formación. Sin embargo, el estudio aquí realizado demuestra afinidad cronológica y biogeográfica de la flora con la fauna anteriormente descrita para esta formación; y ayuda a delimitar el ambiente de depósito, que correspondería a un ambiente deltaico distal (Rosales-Lagarde et al., 2005).
LA FORMACIÓN TUZANCOA Y SU RELACIÓN CON UNIDADES MEXICANAS DE FINALES DEL PALEOZOICO
Los estudios comparativos de la flora del Paleozoico se restringen a Weber (1997) quien comenta de manera general una posible similitud entre la flora fósil de las formaciones Tuzancoa y Matzitzi basado en los dos géneros descritos para la localidad San Mateo 1. Por lo que, los hallazgos de nuevos registros hacen necesaria una nueva comparación entre la paleoflora del Paleozoico reportada para el territorio mexicano. Para México se tiene el reporte de plantas en un total de ocho unidades; sin embargo, para tres de ellas (formaciones Grupera, Santa Rosa Superior e Ixtaltepec) no existen determinaciones taxonómicas o son de un rango superior a género, que no permiten una comparación. Por lo tanto, el siguiente análisis se centra en cinco unidades: la Formación Paso Hondo en el Estado de Chiapas, la Formación Matzizi y el Grupo Patlanoaya en el Estado de Puebla, la Formación Olinalá en el Estado de Guerrero y la Formación Tezuncoa en el Estado de Hidalgo.
Como se observa en la Figura 6, la Formación Tuzancoa presenta una edad de Pensilvánico-Cisuraliense a partir del análisis de la macroflora aquí estudiada. En el caso de la Formación Paso Hondo se ha propuesto una edad de Kunguriense a partir de la presencia de fusulínidos, braquiópodos y ammonites (Hernández-García, 1973), esta fauna junto con la presencia de caliza, lutita y arenisca han permitido proponer un ambiente de depósito de laguna somera con aportes constantes de terrígenos (Hernández-García, 1973; Torres-Martínez et al., 2017). En esta formación solo se tiene una posible determinación y corresponde a un helecho del género Asterotheca; que también está registrado en las otras cuatro unidades. Sin embargo, en la Formación Paso Hondo el ambiente de depósito indica que el material vegetal es alóctono. Lo anterior se ve reflejado en la poca abundancia del material continental a diferencia de lo observado en la Formación Tuzancoa. Por lo que, a partir de los datos hasta el momento publicados se considera una similitud baja entre ambas unidades.
Con respecto a la unidad Patlanoaya, este grupo está dominado por arenisca, lutita, conglomerado y caliza que, junto a los restos de plantas, conodontos, fusulínidos y ammonoideos permiten proponer un origen marino litoral. Con respecto al contenido paleoflorístico, hasta el momento se han reportado un total de siete taxones: Calamites sp., Alethopteris Sternberg, Neuropteris cf. N. schlehami Stur, Sigillaria sp., Supaia White, Pecopteris cyathea Schloteim y Walchia (Villaseñor-Martínez et al., 1987; Silva-Pineda, 2003); a partir de este listado proponen una edad del Misisípico-Cisuraliense. Como se observa en la Tabla 2, comparte con Tuzancoa el 17 % de los géneros; nuevamente esta baja similitud se debe a la diferencia existente entre los ambientes de depósito de ambas formaciones, que sesga el registro paleobotánico del Grupo Patlanoaya y que no permite conjeturar otras causas para esta disimilitud.
Por otro lado, la Formación Matzitzi tiene un amplio rango estratigráfico, con edades que van del Pensilvánico al Triásico de acuerdo con estudios actuales (Silva-Pineda, 1970a; Flores-Barragan, 2019; Juárez-Zúñiga et al., 2020 y Villanueva-Amadoz et al., 2020). Es importante mencionar que esta variación cronológica está relacionada con la localidad en la que se ha realizado el estudio. En este trabajo se utiliza el listado paleoflorístico de la localidad denominada Carretera (Flores-Barragan y Velasco-de León, 2021), a la cual a partir de la macroflora y palinomorfos fósiles se le ha asignado una edad de Pensilvánico (Villanueva-Amadoz et al., 2020) y es la única localidad conocida de la formación Matzitzi parcialmente coetánea con la Formación Tuzancoa.
En la localidad Carretera aflora una sección estratigráfica donde se presenta una sucesión grano decreciente de arenisca de grano mediano a fino de más de 20 metros; la cual ha sido interpretada como planicies de inundación (Centeno-García et al., 2009). En la Tabla 2 se listan los 28 géneros presentes en la formaciones Tuzancoa y Matzitzi (localidad Carretera). Se observa que comparten el 46 % de su diversidad con taxones típicos de finales del Pensilvánico, como Marattiales (Asterotheca y Diplazites Goeppert), Equisetales (Calamites) y Lepidodendrales (Cyperites, Knorria Sternberg, Sigillaria, Stigmaria Brongniart y Syringodendron Sternberg). Esta similitud menor al 50 % refleja una diferencia significativa, probablemente debido al ambiente donde estas comunidades se desarrollaron. En el caso de la Formación Matzitzi las facies propuestas corresponden en general a planicies de inundación (Centeno-García et al., 2009; Bedoya et al., 2021); esta interpretación junto con las adaptaciones presentes en los taxones autóctonos indica un ambiente húmedo, que se apoya en la gran área foliar (hasta 577 cm) presente en algunos taxones (ej. Bjuvia Florin y Velascoa Flores-Barragan y Ortega-Chavez) (Flores-Barragan y Velasco-de León, 2021; Flores-Barragan et al., 2021). Por su parte, el ambiente deltaico distal hasta ahora propuesto para la Formación Tuzancoa, junto con elementos florísticos autóctonos abundantes como Comia, así como la disminución de géneros higrófilos como Calamites, Sigillaria y Asterotheca (Thomas, 1978; Looy, 2007; Phillips y DiMichele, 1992; Taylor et al., 2009; DiMichele et al., 2006; DiMichele et al., 2010; Wilson et al., 2017) indican un clima diferente para ambas unidades y que si bien no son correlacionables permiten dar una visión más general de la vegetación presente a finales del Paleozoico en el territorio nacional.
Por último, con respecto a la Formación Olinalá en el Estado de Guerrero se ha propuesto un ambiente variable de fluvial a margen de rampa, costeros y plataforma externa a partir de la secuencia de conglomerados, limolita, lutita, arenisca y calizas de origen marino (Flores de Dios y Buitrón-Sánchez, 1982; Corona-Esquivel, 1983; Juárez-Arriaga, 2006). También, a partir del ammonoideo Waagenoceras sp. Gemmellaro, de los fusulínidos Polydiexodina sp. Miklukho-Maklay y Codonofusiella extensa Skinner y Wilde y de los bivalvos Nuculavus levatiformis Walcott y Septimyalina burmai Newell se ha propuesto para esta formación una edad de Guadalupiense (Vachard et al., 2004; Juárez-Arriaga, 2006; Guerrero-Sánchez y Quiroz-Barroso, 2013 y Juárez-Arriaga y Murillo-Muñetón, 2020). Para la Formación Olinalá se tienen los registros paleobotánicos de Silva-Pineda et al. (2003), quienes reportan un total de tres géneros, una especie y una afinidad; los cuales corresponden a Asterotheca, Neuropteris sp., Taeniopteris sp., T. fallax Goeppert y cf. Cordaites Unger.
En total, las formaciones Tuzancoa y Olinalá comparten únicamente el 13.6 % de su paleoflora y esta se restringe nuevamente a géneros con una amplia distribución cronológica y ecológica como lo son Asterotheca, Neuropteris sp. y Taeniopteris sp. (Taylor et al., 2009; van Konijnenburg-van Cittert et al., 2017). Si bien las plantas fueron depositadas en ambientes similares, nuevamente parece existir un fuerte sesgo tafonómico y/o falta de estudios en las plantas de la Formación Olinalá que no permite ahondar en la comparación.
Con este análisis comparativo a partir de datos macroflorísticos del Paleozoico de México se observa que, si bien se tienen reportes en ocho unidades distintas, en la mayoría los casos los datos son escasos, con una taxonomía incompleta. También destaca la ausencia de datos de abundancia y riqueza que permitan conocer a profundidad la paleoflora, por lo cual, estudios como el aquí realizado son de suma importancia para comprender mejor la vegetación del Paleozoico superior de México en su correspondiente contexto estratigráfico.

Figura 6. Correlación de las rocas paleozoicas de México con registro paleoflorístico determinado. Modificado de Centeno-García et al. (2009) (Formación Matzitzi), López-Ramos, (1979) (Formación Paso Hondo), Ramos-Arias et al. (2008) y Vachard y Flores de Dios (2002) (Grupo Patlanoaya), Rosales-Lagarde et al. (2005) y Buitrón-Sánchez et al. (2017) (Formación Tuzancoa), Vachard et al. (2004), Juárez-Arriaga (2006), Guerrero-Sánchez y Quiroz-Barroso (2013) y Juárez-Arriaga y Murillo-Muñetón (2020) (Formación Olinalá).
Tabla 2. Tabla de diversidad de los géneros descritos para la Formación Matzitzi, Grupo Patlanoaya, Formación Paso Hondo, Formación Olinalá y Formación Tuzancoa.
|
Orden y/o Familia |
Género |
Puebla |
Chiapas |
Guerrero |
Hidalgo |
|
|
Formación Matzitzi |
Grupo Patlanoaya |
Formación Paso Hondo |
Formación Olinalá |
Formación Tuzancoa |
||
|
Cordaithantales |
cf. cordaites |
* |
||||
|
Cycadales |
Nilssonia Brongniart |
* |
||||
|
Dycranophyllales |
Dycranophyllum Grand'Eury |
* |
||||
|
Equisetales |
Calamites |
* |
* |
* |
||
|
Glossopteridales |
Glossopteris Brongniart |
* |
||||
|
Incertae sedis |
Cordaicarpus |
* |
||||
|
Incertae sedis |
Cornocarpus |
* |
||||
|
Incertae sedis |
Holcospermun Nathorst |
* |
||||
|
Incertae sedis |
Lesleya Lesquereux |
* |
||||
|
Incertae sedis |
Taeniopteris |
* |
* |
* |
||
|
Incertae sedis |
Trigonocarpus Brongniart |
* |
||||
|
Lepidodendrales |
Asolanus |
* |
||||
|
Lepidodendrales |
Cyperites |
* |
* |
|||
|
Lepidodendrales |
Knorria |
* |
* |
|||
|
Lepidodendrales |
Sigillaria |
* |
* |
* |
||
|
Lepidodendrales |
Stigmaria |
* |
* |
|||
|
Lepidodendrales |
Syringodendron |
* |
* |
|||
|
Marattiales |
Astherotheca |
* |
* |
* |
* |
* |
|
Marattiales |
Diplazites |
* |
* |
|||
|
Marattiales |
Lobatopteris |
* |
||||
|
Mariopteridaceae |
Mariopteris |
* |
||||
|
Peltaspermales |
Alethopteris |
* |
||||
|
Peltaspermales |
Comia |
* |
* |
|||
|
Peltaspermales |
cf. Macroneuropteris |
* |
||||
|
Peltaspemales |
cf. Linopteris |
* |
||||
|
Peltaspermales |
Neuropteris |
* |
* |
* |
* |
|
|
Peltaspermales |
Odontopteris |
* |
* |
|||
|
Peltaspermales |
Ovopteris Potonie |
* |
||||
|
Peltaspermales |
Supaia |
* |
||||
|
Voltziales |
Calnalia |
* |
||||
|
Voltziales |
Majonicaceae |
* |
* |
|||
|
Voltziales |
Walchia |
* |
||||
CONCLUSIONES
El hallazgo de una nueva localidad y el aumento en el número de muestreos de las localidades ya conocidas (Calnali 2 y la Virgen 2) permitieron incrementar la diversidad florística conocida de la Formación Tuzancoa, con la descripción de cinco nuevos registros entre estos; estructuras reproductoras de la familia Majonicaceae, que apoya la presencia de este grupo en esta región de Hidalgo. Odontopteris cf. brardii y Mariopteris, del Carbonífero de Euroamérica y España respectivamente. Taeniopteris feddeni del Pérmico de la India y T. tenuis del Carbonífero de Europa. Es importante mencionar que los últimos tres taxones mencionados aumentan su distribución geográfica, siendo en el caso de Mariopteris y Taeniopteris tenuis los reportes más sureños y en Taeniopteris feddeni, el registro más norteño. Estos reportes permiten proponer que la paleoflora aquí descrita tiene una edad de Pensilvánico-Pérmico inferior y con afinidad al reino Euroamericano. También, es importante subrayar que el hallazgo de nuevas localidades permitirá comprender mejor el ambiente de depósito de la formación hasta ahora propuesto; ya que en el caso de La Virgen 2 y San Mateo 2 los especímenes considerados autóctonos indican un ambiente meramente continental. Por otro lado, la comparación con otras unidades con fósiles de plantas demuestra la limitada comprensión que se tiene hasta el momento, impidiendo una comparación adecuada entra la flora de Hidalgo con la del Grupo Patlanoaya y de las formaciones Paso Hondo y Olinalá. La única comparación posible de realizar con los datos actuales corresponde a la formación Matzitzi, concretamente con la localidad Carretera, ambas indican una típica vegetación de finales del Carbonífero; sin embargo, las diferencias taxonómicas demuestran diferentes condiciones ambientales, por lo cual no se consideran correlacionables, pero si complementarias. Lo anteriormente expuesto es un indicador de la importancia de continuar con este tipo de estudios, que permitan dilucidar los procesos evolutivos y biogeográficos que afectaron a la flora mexicana de finales del Paleozoico.
AGRADECIMIENTOS
Los resultados de este trabajo forman parte de la tesis doctoral del primer autor, y da las gracias por el apoyo al Posgrado en Ciencias Biológicas (Posgrado en Ciencias Biológicas, Facultad de Estudios Superiores Zaragoza) de la Universidad Nacional Autónoma de México UNAM y la beca CONACyT 762406. Agradecemos también a E.L. Ortiz-Martínez, E. Ortega-Chávez, P. C. Martínez-Martínez, D.E. Lozano-Carmona y D. Guzmán-Madrid por su ayuda en el desarrollo del trabajo de campo. Deseamos también agradecer al editor en jefe el Dr. Ángel F. Nieto Samaniego sus comentarios y correcciones que ayudaron a mejorar en trabajo. También agradecemos el trabajo realizado por los revisores y muy especialmente la exhaustiva revisión realizada por el Dr. Edgar Juárez Arriaga que resultó muy constructiva contribuyendo a la mejora del trabajo.
REFERENCIAS
Artabe, A.E., 1985, Estudio sistemático de la Tafoflora triásica de Los Menucos, Provincia de Río Negro, Argentina. Parte I, Sphenophyta, Filicophyta y Pteridospermophyta: Ameghiniana, 22, 3-22.
Bedoya, A., Anaya-Guarneros, J.A., Abdullin, F., Martini, M., Solari, L., 2021, Provenance analysis of the Matzitzi and Agua de Mezquite formations, southern Mexico: Different fluvial successions formed during late Paleozoic and post-Middle Jurassic time along the southernmost North America Pacific margin: Journal of South American Earth Sciences, 105, https://doi.org/10.1016/j.jsames.2020.102999
Bose, M.N., Taylor, E.L., Taylor, N.T., 1990, Gondwana Floras of India and Antarctica-A Survey and Appraisal, en Taylor, N.T. (ed.), Antarctic Paleobiology: New York, NY, Springer-Verlag, 118-148, https://doi.org/10.1007/978-1-4612-3238-4_10
Brongniart, A., 1822, Sur la classification et la distribution des végétaux fossiles: Mémoires du Muséum National d'Histoire naturelle, 8, 203-240, 297-348.
Brongniart, A., 1828, Prodrome d’une Histoire des végétaux fossiles: Dictionnaire des sciences naturelles: Francia, Forgotten Books, 223 pp.
Buitrón-Sánchez, B.E., López-Lara, O., Vachard, D., Hernández Barroso, S., 2017, Algunos crinoides (Echinodermata-Crinoidea) del Pérmico de la región de Pemuxco, Hidalgo: Boletín de la Sociedad Geológica Mexicana, 69(1), 21-34.
Carrillo-Bravo, J., 1965, Estudio Geológico de una parte del Anticlinorio de Huayacocotla: Boletín de la Asociación Mexicana de Geólogos Petroleros, 17(5-6), 73-96.
Centeno-García, E., 2005, Review of Upper Paleozoic and Lower Mesozoic stratigraphy and depositional environments of central and West Mexico: Constraints on terrane analysis and paleogeography, en Anderson, T.H., Nourse, J.A., McKee, J.W., Steiner, M.B. (eds.), The Mojave-Sonora megashear hypothesis: Development, assessment and alternatives: Geological Society of America Special Paper, 393, 233-258.
Centeno-García, E., Mendoza-Rosales, C.C., Silva-Romo, G., 2009, Sedimentología de la Formación Matzitzi (Paleozoico superior) y significado de sus componentes volcánicos, región de Los Reyes Metzontla-San Luis Atolotitlán, Estado de Puebla: Revista Mexicana de Ciencias Geológicas, 26(1), 18-36.
Cleal, C.J., Shute, C.H., 1991, Proposal to conserve Odontopteris (Brongniart) Sternberg (Fossils) against Odontopteris Bernhardi (Schizaeaceae): Taxon, 40, 130-132.
Clement-Westerhof, J.A., 1987, Aspects of Permian palaeobotany and palynology; VII, The Majonicaceae, a new family of Late Permian conifers: Review Palaeobotany Palynology, 52, 375-402.
Corona-Esquivel, R., 1983, Estratigrafía de la región de Olinalá-Tecocoyunca, Noreste del Estado de Guerrero: Revista del Instituto de Geología, 5(1), 17-24.
DiMichele, W.A., Tabor, N.J., Chaney, D., Nelson, W.J., 2006, From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras: Geological Society of America, 399, 223-248.
DiMichele, W.A., Cecil, C.B., Montañez, I.P., Falcon-Lang, H.J., 2010, Cyclic changes in Pennsylvanian paleoclimate and effects on floristic dynamics in tropical Pangaea: International Journal of Coal Geology, 83, 329-344.
Doubinger, J., Vetter, P., 1959, Contribution a l´étude du genre Taeniopteris Brongniart: Bulletin de la Société Géologique de France, S7-I (9), 932-938.
Flores-Barragan, M.A., 2019, Las Ginkgophytas de la Formación Matzitzi, implicaciones taxonómicas y ecológicas: México, Universidad Nacional Autónoma de México, Tesis de maestría, 93 pp.
Flores-Barragan, M.A., Velasco-de León, M.P., 2021, New records of Bjuvia and Nilssonia from the Permian of Mexico: Palaeontologia Electronica, 24(3), a28, https://doi.org/10.26879/1109
Flores-Barragan, M.A., Velasco-de León, M.P., Ortega-Chavez, E., 2021, New genus megaphyllous leaves from the Upper Paleozoic of Mexico Velascoa pueblensis gen. Nov: Journal of South American Earth Sciences, 110, https://doi.org/10.1016/j.jsames.2021.103408
Flores de Dios, A., Buitrón-Sánchez, B.E., 1982, Revisión y aportes a la estratigrafía de la Montaña de Guerrero: Universidad Autónoma de Guerrero, Serie Técnico Científica, (12), 1-28.
Forte, G., Kustatscher, E., van Konijnenburg-van Cittert, J.H.A., Looy, C.V., Kerp, H., 2017, Conifer diversity in the Kungurian of Europe-Evidence from dwarf-shoot morphology: Review of Palaeobotany and Palynology, 244, 308-315.
Gastaldo, R.A., 1988, Conspectus of phytotaphonomy en DiMichele, W.A., Wing, S.L., (eds.), Methods and Applications of Plant Paleoecology: Notes for a Short Course: Paleontological Society Special Publication: Inglaterra, Cambridge University Press, 14-28.
Gastaldo, R.A., Boersma, M., 1983, A reinvestigation of Early Pennsylvanian species of Mariopteris from the Appalachian region. I. Karinopteris, Mariopteris and the "pottsvillea complex": Review of Palaeobotany and Palynology, 38, 185-226, https://doi.org/10.1016/0034-6667(83)90023-4
Greenwood, D.R., 1991, The taphonomy of plants macrofossils, en S.K., Donovan (ed.), The processes of fossilization: Londres, Belhaven Press, 141-169.
Guerrero-Sánchez, M., Quiroz-Barroso, S.A., 2013, Bivalvos guadalupianos (Pérmico medio) de la Formación Olinalá, Guerrero, México: Boletín de la Sociedad Geológica Mexicana, 65 (3), 533-551.
Gutiérrez-Alejandre, J.F., 2018, Contribución al conocimiento geológico del Anticlinorio de Huayacocotla, mediante el análisis sedimentológico-estratigráfico de afloramientos del Paleozoico superior, en la región de Calnali, Hgo: México, Universidad Nacional Autónoma de México, Tesis de licenciatura, 73 pp.
Hernández-Castillo, G.R., Silva-Pineda, A., Cevallos Ferriz, S.R.S., 2014, Early Permian conifer remains from Central Mexico and reevaluation of Paleozoic conifer morphotaxa: Boletín de la Sociedad Geológica Mexicana, 66, 85-96.
Hernández-García, R., 1973, Paleogeografía del Paleozoico de Chiapas, México: Boletín de la Asociación de Geólogos Petroleros, 25 (1-3), 77-134.
Juárez-Arriaga, E., 2006, Marco de Estratigrafía de Secuencias para la Sucesión Sedimentaria Terrígeno-Carbonatada del Paleozoico Superior del área de Olinalá, Guerrero, México: México, Universidad Nacional Autónoma de México Tesis de maestría, 93 pp.
Juárez-Arriaga, E., Murillo-Muñetón, G., 2020, Arquitectura estratigráfica, ambientes de depósito y geocronología de la Formación Olinalá (Pérmico tardío), noreste de Guerrero, México: Revista Mexicana de Ciencias Geológicas, 37(3), 179-196.
Juárez-Zúñiga, S., Solari., L.A., Ortega-Obregón, C., 2020, Permian igneous clasts from the Matzitzi Formation, southern Mexico: isotopic constraints on the final amalgamation of Pangea: Geological Society of London, Publicación especial, https://doi.org/10.1144/SP503-2019-238
Knight, J., 1983, The stratigraphy of the Stephanian rocks of the Sabero Coalfield, León (NW Spain) and an investigation of the fossil flora: Palaeontographica, B187, 155-248.
Looy, C.V., 2007, Extending the range of derived late Paleozoic conifers: Lebowskia gen. nov. (majonicaceae): International Journal of Plant Sciences, 168(6), 957-972.
López-Ramos, E., 1979, Geología de México: México, Ediciones Escolares, tomo 3, 446 pp.
Miall, D., 2006, The geology of fluvial deposits sedimentary facies, basin Analysis, and petroleum geology, 4 edición: Berlin, Alemania, Springer, Berlin, Heidelberg, 582 pp.
Navarro-Santillán, D., Sour-Tovar, F., Centeno-García, E., 2002, Lower Mississippian (Osagean) brachiopods from the Santiago Formation, Oaxaca, México: Stratigraphic and tectonic implications: Journal of South American Earth Sciences, 15, 327-336.
Němejc, F., 1968, Paleobotanika III. Systematická část. Rostliny nahosemenné: Praga, Republica Checa, Nakladatelství Československé Akademie, 478 pp.
Ortega-Gutiérrez, F., Ruiz, J., Centeno-García, E., 1995, Oaxaca, a Proterozoic microcontinent accreted to North America during the late Paleozoic: Geology, 23, 1127-1130.
Ortiz-Martínez, E.L., Velasco-de León, M.P., 2019, First report on seeds dispersed in Tuzancoa Formation, Hidalgo: International Journal of Innovative Studies in Science and Enginnering Technology, 5, 1-5.
Phillips, T.L., DiMichele, W.A., 1992, Comparative Ecology and Life-History Biology of Arborescent Lycopsids in Late Carboniferous Swamps of Euramerica: Annals of the Missouri Botanical Garden, 79 (3), 560-588.
Quiroz-Barroso, S.A., Pojeta, J., Sour-Tovar, F., Morales-Soto, S., 2000, Pseudomulceodens, a Mississippian Rostroconch from Mexico: Journal of Paleontology, 74(6), 1184-1186.
Quiroz-Barroso, S., Sour-Tovar, F., Centeno-García, E., 2012, Gasteropodos y bivalvos cisuralianos (Pérmico inferior) de Otlamalacatla, Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 29(1), 158-178.
Ramos-Arias, M.A., Keppie, J.D., Ortega-Rivera, A., Lee, J.W.K., 2008, Extensional Late Paleozoic deformation on the western margin of Pangea, Patlanoaya area, Acatlán Complex, southern Mexico: Tectonophysics, 448, 60-76.
Remy, W., Remy, R., 1975, Beiträge zur Kenntnis des Morpho-Genus Taeniopteris Brongniart: Argumenta Paleobotánica, 4, 31-37.
Rosales-Lagarde, L., Centeno-García, E., Dostal, J., Sour-Tovar, F., Ochoa-Camarillo, H., Quiroz-Barroso, S., 2005, The Tuzancoa Formation: Evidence of an Early Permian submarine continental arc in east-central Mexico: International Geology Review, 47(9), 901-919.
Silva-Pineda, A., 1963, Plantas del Triásico superior del estado de Hidalgo: Paleontología Mexicana, 18, 1-12.
Silva-Pineda, A., 1970a, Plantas del Pensilvánico de la región de Tehuacán: Paleontología Mexicana, 29, 1-108.
Silva-Pineda, A., 1970b, Fructificación de pteridosperma en el Pensilvánico de Oaxaca (resumen), en Primera Convención Nacional de la Sociedad Geológica Mexicana: Ciudad de México, México, Sociedad Geológica Mexicana, 85-86.
Silva-Pineda, A., 1984, Revisión Taxonómica y tipificación de las plantas Jurásicas colectadas y estudiadas por Wieland (1914) en la región de El Consuelo, Oaxaca: Paleontología Mexicana, 49, 1-104.
Silva-Pineda, A., 1987, Algunos elementos paleoflorísticos del Pérmico de la región de Calnali, Estado de Hidalgo: Paleontología Mexicana, 1(l), 313-327.
Silva-Pineda, A., 2003, Flora del Pérmico de Izúcar de Matamoros, Puebla, en Soto, L.A. (ed.), Ayala-Castañares: universitario impulsor de la investigación científica: México, Instituto de Ciencias del Mar y Limnología UNAM, 303-309.
Silva-Pineda, A., Buitrón-Sánchez, B.E., 1998, Paleontología de México. Plantas vasculares fósiles: Ciudad de México, México, Facultad de Ingeniería, Universidad Nacional Autónoma de México, 93 pp.
Silva-Pineda, A., Buitrón-Sánchez, B.E., Arellano-Gil, J., Vachard, D., Ramírez, J., 2003, Permian Continental and Marine Biota of South-Central México A Synthesis en Bartolini, C., Buffler, R.T., Blickwede, J., (eds.), The Circum-Gulf of Mexico and the Caribbean: Hydrocarbon habitats, basin formation, and plate tectonics: American Association of Petroleum Geologist Memoir, 79, 462-275.
Sour-Tovar, F., Pérez-Huerta, A., Quiroz-Barroso, S.A., Centeno-García, E., 2005, Braquiópodos y trilobite del Pérmico Inferior del noroeste del Estado de Hidalgo, México: Revista Mexicana de Ciencias Geológicas 22, 24-35.
Sternberg, K.V., 1825, Versuch einer geognostisch- botanischen Darstellung der Flora der Vorwelt: Leipzig, Alemania, Kommission im Deutschen Museum, 220 pp.
Stewart, W.N., Rothwell, G.W., 1993, Paleobotany and the evolution of plants: Cambridge, Reino Unido, Cambridge University Press, 536 pp.
Taylor, T.N., 1981, Paleobotany. An introduction to fossil plant biology: New York, McGraw-Hill book co. 589 pp.
Taylor, T.N., Taylor, E.L., Krings, M., 2009, Paleobotany the biology and evolution of fossil plants: USA, Elsevier Inc., 2nd Edition 1230 pp.
Thomas, B.A., 1978, Carboniferous Lepidodendraceae and Lepidocarpacea: Botanical Review, 44 (3), 321-364.
Torres-Martínez, M.A., Barragán, R., Sour-Tovar, F., González-Mora, S., 2017, Depositional paleoenvironments of the Lower Permian (upper Cisuralian) carbonate succession of Paso Hondo Formation in Chiapas State, southeastern Mexico: Journal of South American Earth Sciences, 79, 254-263.
Vachard, D., Flores de Dios, A., 2002, Discovery of latest Devonian/earliest Mississippian microfossils in San Salvador Patlanoaya (Puebla, Mexico); biogeographic and geodynamic consequences: Geoscience, 334, 1095-1101.
Vachard, D., Pantoja, J., 1997, Evolution of Mexico during the Upper Paleozoic, en Segunda Convención sobre la evolución geológica de México y recursos asociados: Pachuca México, México, Universidad Autónoma de Hidalgo, Instituto de Investigaciones en Ciencias de la Tierra; Universidad Nacional Autónoma de México, Instituto de Geología, 49-53.
Vachard, D., Flores-de Dios, D., Pantoja, J., Buitron-Sanchez, B.E., Arellano, J., Grajales, M., 2000, Les fusulines du Mexique, une revue biostratigraphique et paléogéographique: Geobios, 33, 655-679.
Vachard, D., Flores de Dios, A., Buitrón-Sánchez, B.E., 2004, Guadalupian and Lopingian (Middle and Late Permian) deposits from Mexico and Guatemala, a review with new data: Geobios, 37, 99-115.
van Konijnenburg-van Cittert, J.H.A., Pott, C., Cleal, C.J., Zijlstra, G., 2017, Differentiation of the fossil leaves assigned to Taeniopteris, Nilssoniopteris and Nilssonia with a comparison to similar genera: Review of Palaeobotany and Palynology, 237, 100-106, https://doi.org/10.1016/j.revpalbo.2016.11.009
Velasco-de León, M.P., Ortiz-Martínez, E.L., 2010, Nuevas especies de Quercus (Fagaceae) en el Plioceno de Santa María Amajac, Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 27, 264-277.
Velasco-de León, M.P., Ortiz Martínez, E.L., Lozano-Carmona, D. E., Flores-Barragan, M.A., Arellano, G.J., Santillán N.P., 2020, Paleofloristic diversity and paleoenvironmental interpretation of new Cisuralian localities in the south-central sector of the Anticlinorium of Huayacocotla, Hidalgo, Mexico: Geobios, 63, 53-66. https://doi.org/10.1016/j.geobios.2020.07.004
Villanueva-Amadoz, U., Navarroa, M. G., Juncal, M. A., Diez, J. B., 2020, Paleobotanical and palynological evidence for the age of the Matzitzi Formation, Mexico: Comptes Rendus Géoscience-Sciences de la Planète, 352, 515-538.
Villaseñor-Martínez, A.B., Martínez-Cortés, A.M., Contreras y Montero, B., 1987, Bioestratigrafía del Paleozoico Superior de San Salvador Patlanoaya, Puebla, México: Revista de la Sociedad Mexicana de Paleontología, 1, 396-417.
Wagner, R.H., 1959, Some Stephanian Pecopteris from N.W. Spain: Mededelingen van de Geologische Stichting, Nieuwe Serie, 12, 5-23.
Weber, R., 1997, How Old Is the Triassic Flora of Sonora and Tamaulipas and News on Leonardian Floras in Puebla and Hidalgo, Mexico: Revista Mexicana de Ciencias Geológicas, 14, 225-243.
Wilson, J.P., Montañez, I.P., White, J.D., DiMichele, W.A., McElwain, J.C., Poulsen, C.J., Hren, T., 2017, Dynamic Carboniferous tropical forests: new views of plant function and potential for physiological forcing of climate: New phytologist, 215, 1333-1353.
Zeiller, M.R., 1879, Presentation de l'Atlas du Tome IV de l'Explication de la Carte G6ologique. de la France, et note sur le genre Mariopteris: Bulletin de la Société Géologique de France, 7, 92-99.
Zeiller, M.R., 1882, Examen de la flore fossile des couches de charbon du Tong-King: Annales des Mines, 8, 299-352.